

The NH2 tail of the novel histone variant H2BFWT exhibits properties distinct from conventional H2B with respect to the assembly of mitotic chromosomes
- PMID: 16449661
- PMCID: PMC1367197
- DOI: 10.1128/MCB.26.4.1518-1526.2006
The NH2 tail of the novel histone variant H2BFWT exhibits properties distinct from conventional H2B with respect to the assembly of mitotic chromosomes
Abstract
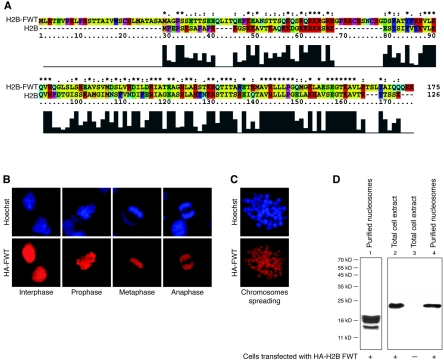
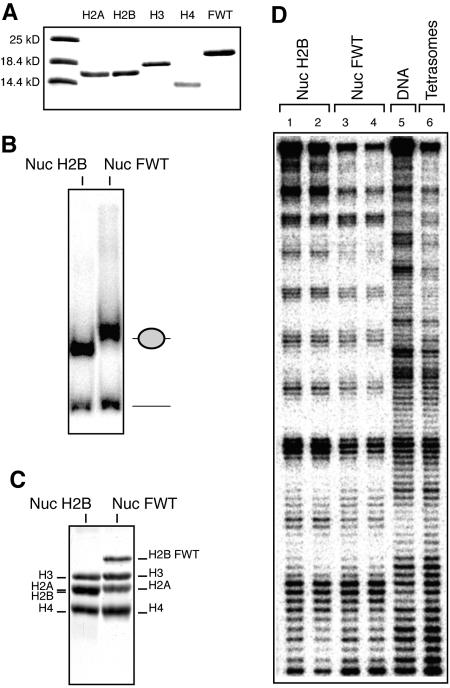
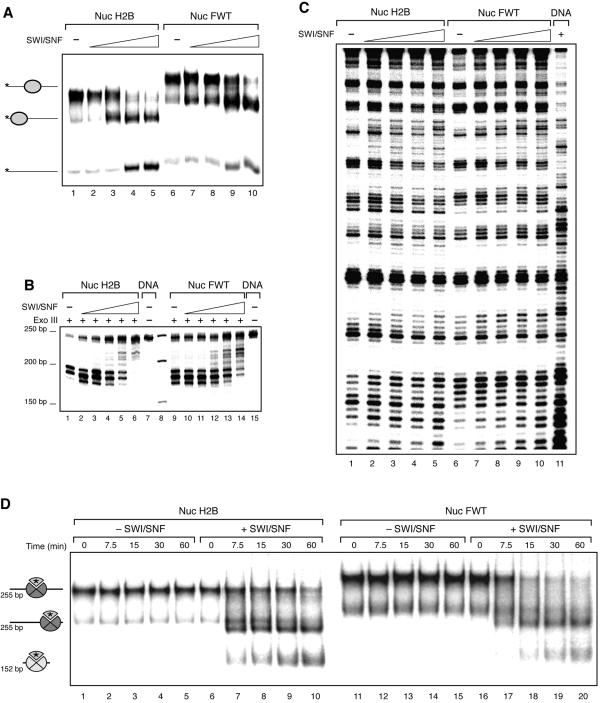
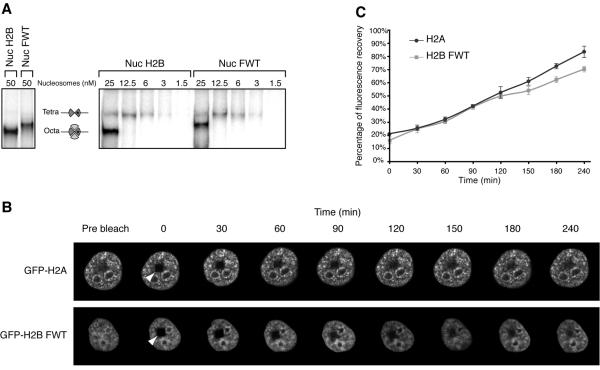
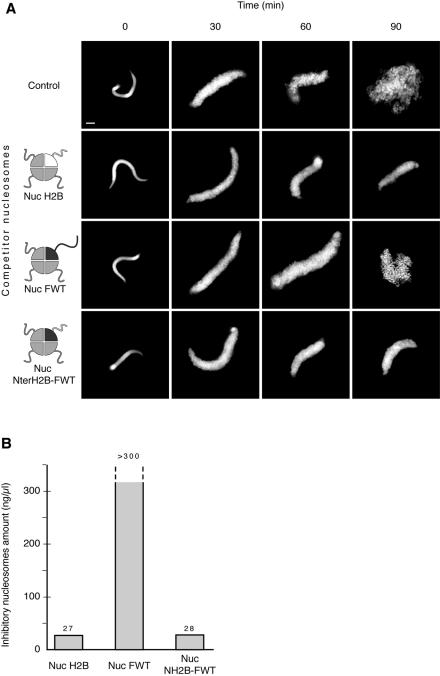
We have studied the functional and structural properties of nucleosomes reconstituted with H2BFWT, a recently identified putative histone variant of the H2B family with totally unknown function. We show that H2BFWT can replace the conventional histone H2B in the nucleosome. The presence of H2BFWT did not affect the overall structure of the nucleosome, and the H2BFWT nucleosomes exhibited the same stability as conventional nucleosomes. SWI/SNF was able to efficiently remodel and mobilize the H2BFWT nucleosomes. Importantly, H2BFWT, in contrast to conventional H2B, was unable to recruit chromosome condensation factors and to participate in the assembly of mitotic chromosomes. This was determined by the highly divergent (compared to conventional H2B) NH2 tail of H2BFWT. These data, in combination with the observations that H2BFWT was found by others in the sperm nuclei and appeared to be associated with the telomeric chromatin, suggest that H2BFWT could act as a specific epigenetic marker.
Figures
References
-
- An, W., V. B. Palhan, M. A. Karymov, S. H. Leuba, and R. G. Roeder. 2002. Selective requirements for histone H3 and H4 N termini in p300-dependent transcriptional activation from chromatin. Mol. Cell 9:811-821. - PubMed
-
- Angelov, D., A. Molla, P. Y. Perche, F. Hans, J. Cote, S. Khochbin, P. Bouvet, and S. Dimitrov. 2003. The Histone variant macroH2A interferes with transcription factor binding and SWI/SNF nucleosome remodeling. Mol. Cell 11:1033-1041. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases