

A functional module of yeast mediator that governs the dynamic range of heat-shock gene expression
- PMID: 16452140
- PMCID: PMC1456402
- DOI: 10.1534/genetics.105.052738
A functional module of yeast mediator that governs the dynamic range of heat-shock gene expression
Abstract
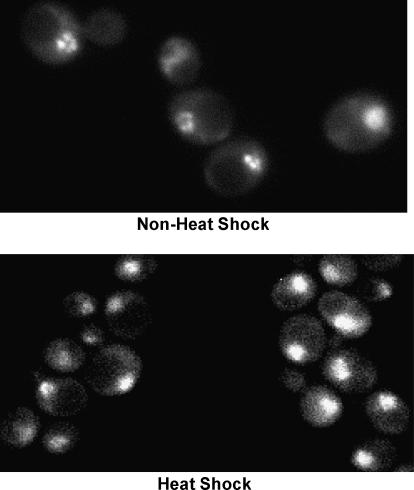
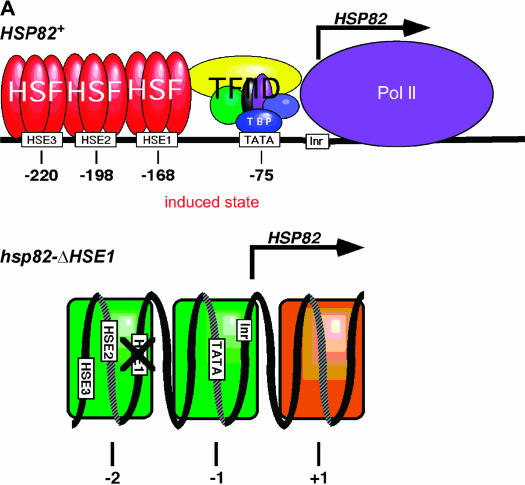
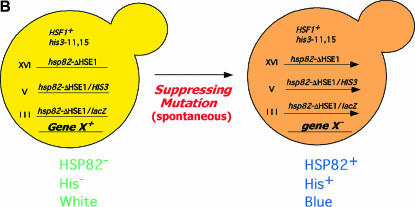
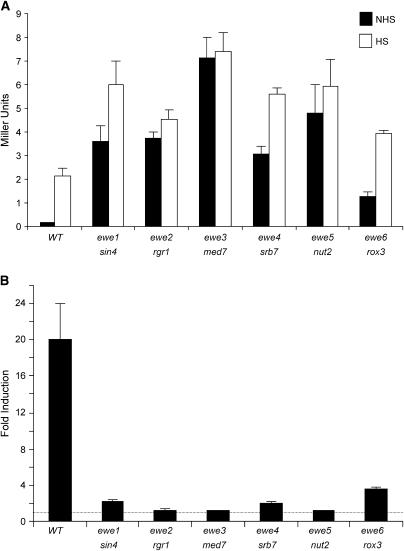
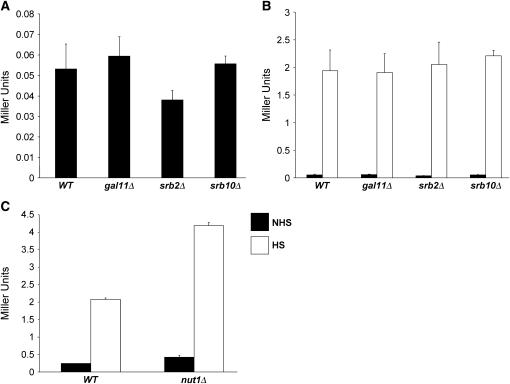
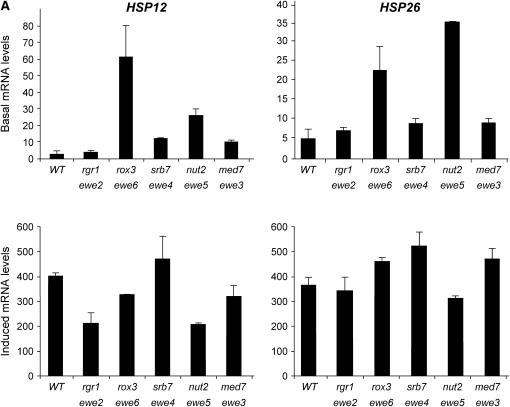
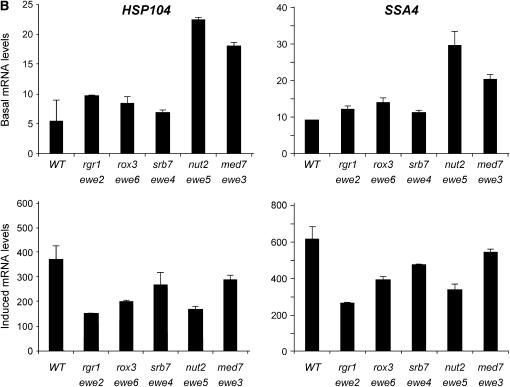
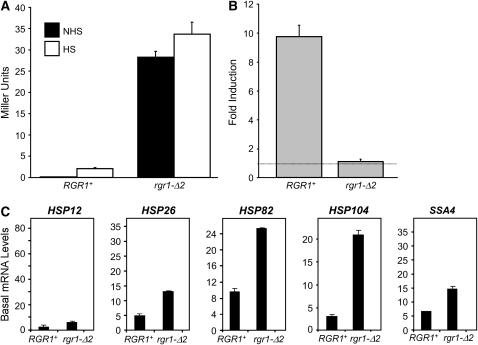
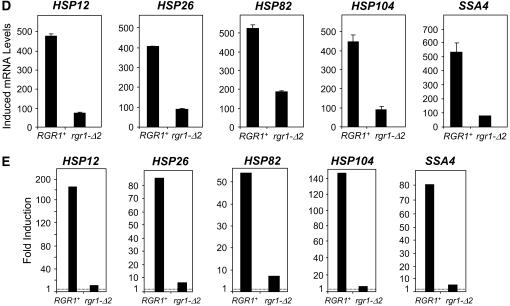
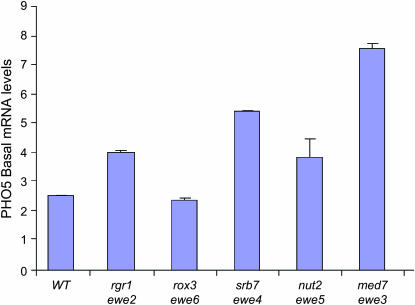
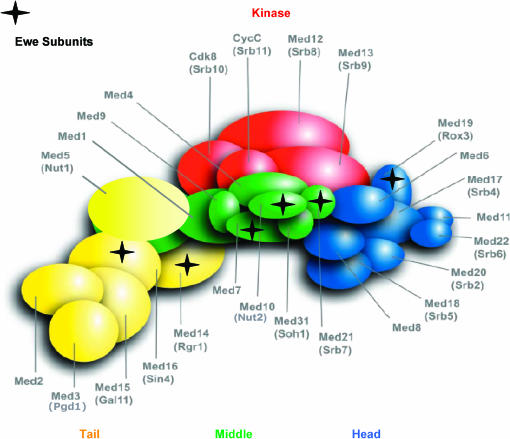
We report the results of a genetic screen designed to identify transcriptional coregulators of yeast heat-shock factor (HSF). This sequence-specific activator is required to stimulate both basal and induced transcription; however, the identity of factors that collaborate with HSF in governing noninduced heat-shock gene expression is unknown. In an effort to identify these factors, we isolated spontaneous extragenic suppressors of hsp82-deltaHSE1, an allele of HSP82 that bears a 32-bp deletion of its high-affinity HSF-binding site, yet retains its two low-affinity HSF sites. Nearly 200 suppressors of the null phenotype of hsp82-deltaHSE1 were isolated and characterized, and they sorted into six expression without heat-shock element (EWE) complementation groups. Strikingly, all six groups contain alleles of genes that encode subunits of Mediator. Three of the six subunits, Med7, Med10/Nut2, and Med21/Srb7, map to Mediator's middle domain; two subunits, Med14/Rgr1 and Med16/Sin4, to its tail domain; and one subunit, Med19/Rox3, to its head domain. Mutations in genes encoding these factors enhance not only the basal transcription of hsp82-deltaHSE1, but also that of wild-type heat-shock genes. In contrast to their effect on basal transcription, the more severe ewe mutations strongly reduce activated transcription, drastically diminishing the dynamic range of heat-shock gene expression. Notably, targeted deletion of other Mediator subunits, including the negative regulators Cdk8/Srb10, Med5/Nut1, and Med15/Gal11 fail to derepress hsp82-deltaHSE1. Taken together, our data suggest that the Ewe subunits constitute a distinct functional module within Mediator that modulates both basal and induced heat-shock gene transcription.
Figures
Similar articles
-
Role of Mediator in regulating Pol II elongation and nucleosome displacement in Saccharomyces cerevisiae.Genetics. 2012 May;191(1):95-106. doi: 10.1534/genetics.111.135806. Epub 2012 Feb 29. Genetics. 2012. PMID: 22377631 Free PMC article.
-
Cooperative binding of heat shock factor to the yeast HSP82 promoter in vivo and in vitro.Mol Cell Biol. 1999 Mar;19(3):1627-39. doi: 10.1128/MCB.19.3.1627. Mol Cell Biol. 1999. PMID: 10022851 Free PMC article.
-
Functional and physical interactions within the middle domain of the yeast mediator.Mol Genet Genomics. 2006 Aug;276(2):197-210. doi: 10.1007/s00438-006-0135-7. Epub 2006 Jun 7. Mol Genet Genomics. 2006. PMID: 16758199
-
Mediator recruitment to heat shock genes requires dual Hsf1 activation domains and mediator tail subunits Med15 and Med16.J Biol Chem. 2013 Apr 26;288(17):12197-213. doi: 10.1074/jbc.M112.449553. Epub 2013 Feb 27. J Biol Chem. 2013. PMID: 23447536 Free PMC article.
-
The yeast Mediator complex and its regulation.Trends Biochem Sci. 2005 May;30(5):240-4. doi: 10.1016/j.tibs.2005.03.008. Trends Biochem Sci. 2005. PMID: 15896741 Review.
Cited by
-
Histone modifications influence mediator interactions with chromatin.Nucleic Acids Res. 2011 Oct;39(19):8342-54. doi: 10.1093/nar/gkr551. Epub 2011 Jul 8. Nucleic Acids Res. 2011. PMID: 21742760 Free PMC article.
-
Heat shock factor 1 counteracts epigenetic silencing of nuclear transgenes in Chlamydomonas reinhardtii.Nucleic Acids Res. 2013 May 1;41(10):5273-89. doi: 10.1093/nar/gkt224. Epub 2013 Apr 12. Nucleic Acids Res. 2013. PMID: 23585280 Free PMC article.
-
Role of Mediator in regulating Pol II elongation and nucleosome displacement in Saccharomyces cerevisiae.Genetics. 2012 May;191(1):95-106. doi: 10.1534/genetics.111.135806. Epub 2012 Feb 29. Genetics. 2012. PMID: 22377631 Free PMC article.
-
mRNA quality control is bypassed for immediate export of stress-responsive transcripts.Nature. 2016 Dec 22;540(7634):593-596. doi: 10.1038/nature20572. Epub 2016 Dec 12. Nature. 2016. PMID: 27951587
-
Facilitated assembly of the preinitiation complex by separated tail and head/middle modules of the mediator.J Mol Biol. 2012 Jan 20;415(3):464-74. doi: 10.1016/j.jmb.2011.11.020. Epub 2011 Nov 23. J Mol Biol. 2012. PMID: 22137896 Free PMC article.
References
-
- Amoros, M., and F. Estruch, 2001. Hsf1p and Msn2/4p cooperate in the expression of the Saccharomyces cerevisiae genes HSP26 and HSP104 in a gene- and stress type-dependent manner. Mol. Microbiol. 39: 1523–1532. - PubMed
-
- Apone, L. M., C. A. Virbasius, F. C. P. Holstege, J. Wang, R. A. Young et al., 1998. Broad, but not universal, transcriptional requirement for yTAFII17, a histone H3-like TAFII present in TFIID and SAGA. Mol. Cell 2: 653–661. - PubMed
-
- Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman et al., 1995. Current Protocols in Molecular Biology. John Wiley & Sons, New York.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
