

A mutation in Tac1p, a transcription factor regulating CDR1 and CDR2, is coupled with loss of heterozygosity at chromosome 5 to mediate antifungal resistance in Candida albicans
- PMID: 16452151
- PMCID: PMC1456413
- DOI: 10.1534/genetics.105.054767
A mutation in Tac1p, a transcription factor regulating CDR1 and CDR2, is coupled with loss of heterozygosity at chromosome 5 to mediate antifungal resistance in Candida albicans
Abstract
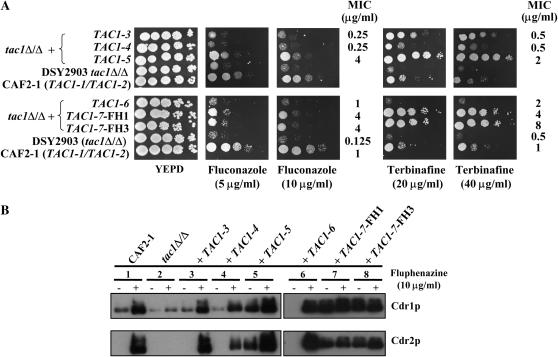
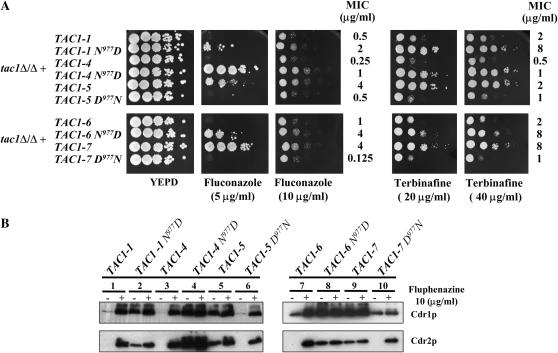
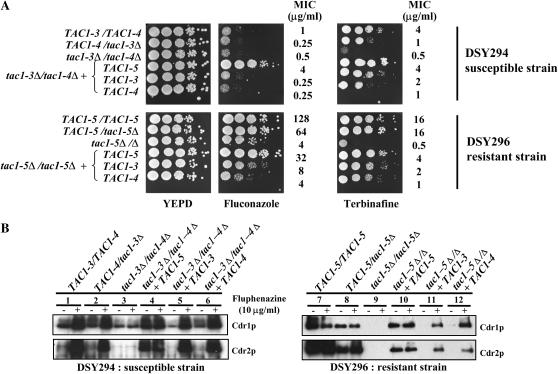
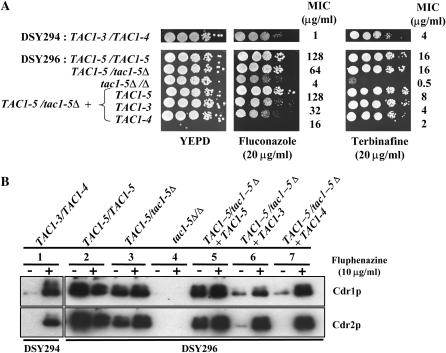
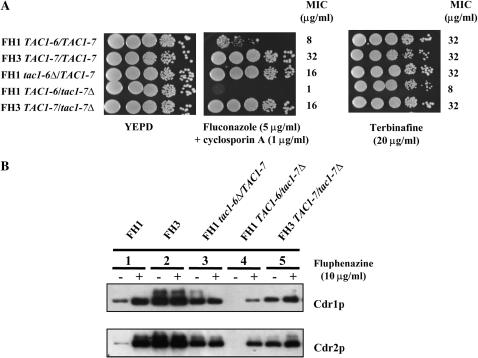
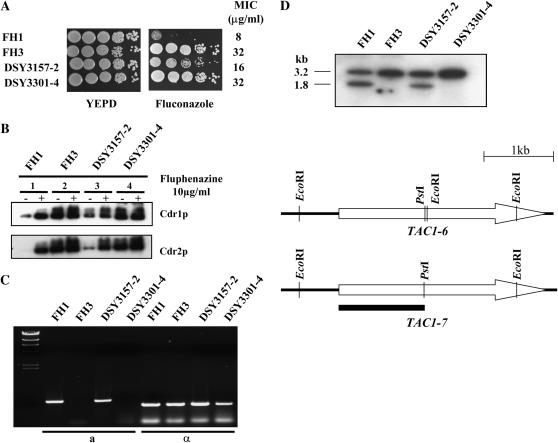
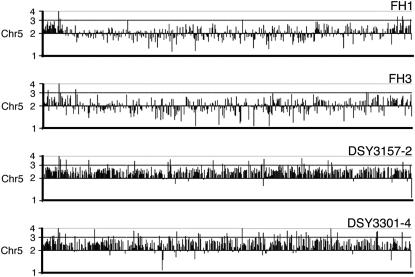
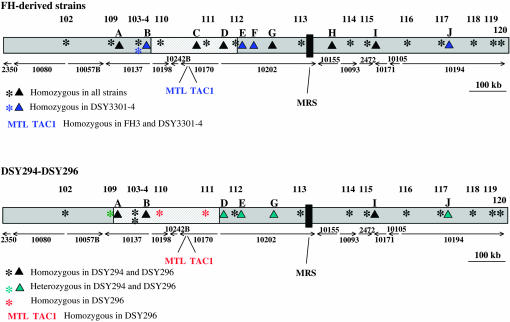
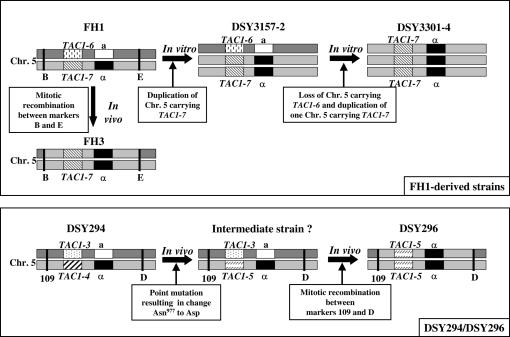
TAC1, a Candida albicans transcription factor situated near the mating-type locus on chromosome 5, is necessary for the upregulation of the ABC-transporter genes CDR1 and CDR2, which mediate azole resistance. We showed previously the existence of both wild-type and hyperactive TAC1 alleles. Wild-type alleles mediate upregulation of CDR1 and CDR2 upon exposure to inducers such as fluphenazine, while hyperactive alleles result in constitutive high expression of CDR1 and CDR2. Here we recovered TAC1 alleles from two pairs of matched azole-susceptible (DSY294; FH1: heterozygous at mating-type locus) and azole-resistant isolates (DSY296; FH3: homozygous at mating-type locus). Two different TAC1 wild-type alleles were recovered from DSY294 (TAC1-3 and TAC1-4) while a single hyperactive allele (TAC1-5) was isolated from DSY296. A single amino acid (aa) difference between TAC1-4 and TAC1-5 (Asn977 to Asp or N977D) was observed in a region corresponding to the predicted activation domain of Tac1p. Two TAC1 alleles were recovered from FH1 (TAC1-6 and TAC1-7) and a single hyperactive allele (TAC1-7) was recovered from FH3. The N977D change was seen in TAC1-7 in addition to several other aa differences. The importance of N977D in conferring hyperactivity to TAC1 was confirmed by site-directed mutagenesis. Both hyperactive alleles TAC1-5 and TAC1-7 were codominant with wild-type alleles and conferred hyperactive phenotypes only when homozygous. The mechanisms by which hyperactive alleles become homozygous was addressed by comparative genome hybridization and single nucleotide polymorphism arrays and indicated that loss of TAC1 heterozygosity can occur by recombination between portions of chromosome 5 or by chromosome 5 duplication.
Figures
References
-
- Carvajal, E., H. B. van den Hazel, A. Cybularz-Kolaczkowska, E. Balzi and A. Goffeau, 1997. Molecular and phenotypic characterization of yeast PDR1 mutants that show hyperactive transcription of various ABC multidrug transporter genes. Mol. Gen. Genet. 256: 406–415. - PubMed
-
- Chambers, S. P., S. E. Prior, D. A. Barstow and N. P. Minton, 1988. The pMTL nic- cloning vectors. I. Improved pUC polylinker regions to facilitate the use of sonicated DNA for nucleotide sequencing. Gene 68: 139–149. - PubMed
-
- Chen, X., B. B. Magee, D. Dawson, P. T. Magee and C. A. Kumamoto, 2004. Chromosome 1 trisomy compromises the virulence of Candida albicans. Mol. Microbiol. 51: 551–565. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
