

Increased levels of the FoxM1 transcription factor accelerate development and progression of prostate carcinomas in both TRAMP and LADY transgenic mice
- PMID: 16452231
- PMCID: PMC1363687
- DOI: 10.1158/0008-5472.CAN-05-3138
Increased levels of the FoxM1 transcription factor accelerate development and progression of prostate carcinomas in both TRAMP and LADY transgenic mice
Abstract
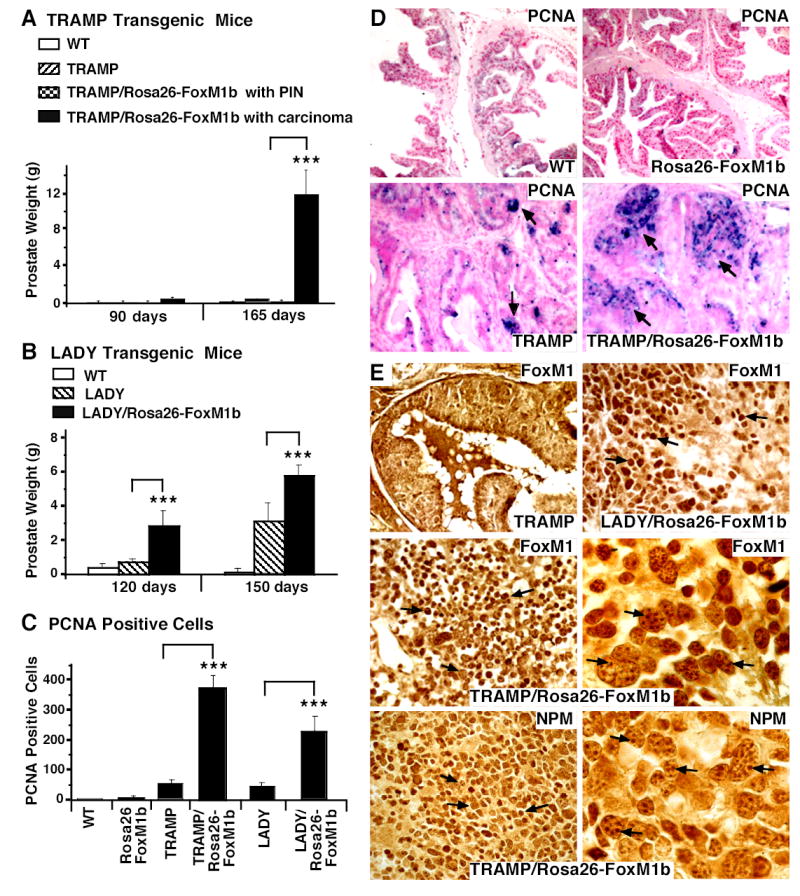
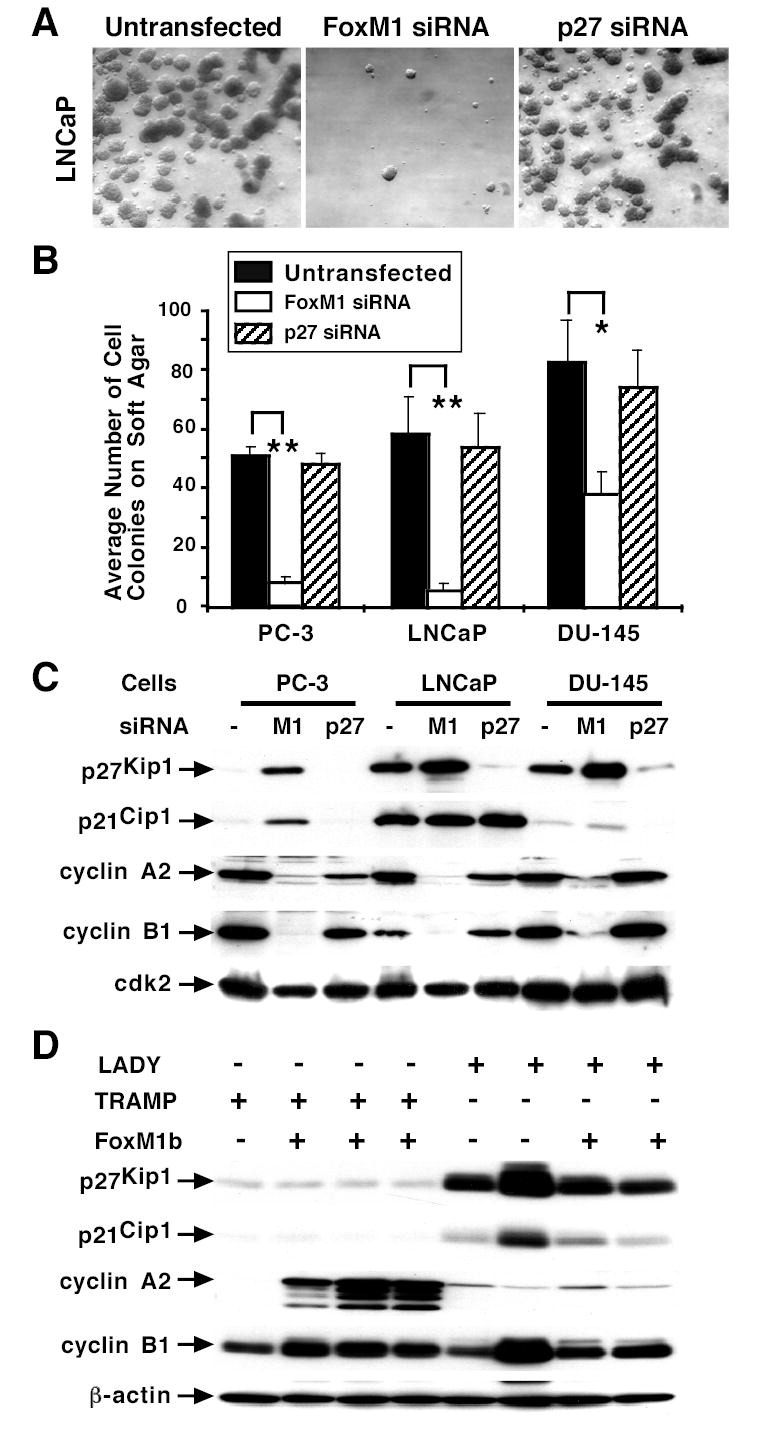
The proliferation-specific Forkhead Box M1 (FoxM1 or FoxM1b) transcription factor is overexpressed in a number of aggressive human carcinomas. Mouse hepatocytes deficient in FoxM1 fail to proliferate and are highly resistant to developing carcinogen-induced liver tumors. We previously developed a transgenic (TG) mouse line in which the ubiquitous Rosa26 promoter was used to drive expression of the human FoxM1b cDNA transgene in all mouse cell types. To investigate the role of FoxM1b in prostate cancer progression, we bred Rosa26-FoxM1b mice with both TRAMP and LADY TG mouse models of prostate cancer. We show that increased expression of FoxM1b accelerated development, proliferation, and growth of prostatic tumors in both TRAMP and LADY double TG mice. Furthermore, development of prostate carcinomas in TRAMP/Rosa26-FoxM1b double TG mice required high levels of FoxM1 protein to overcome sustained expression of the alternative reading frame tumor suppressor, a potent inhibitor of FoxM1 transcriptional activity. Depletion of FoxM1 levels in prostate cancer cell lines PC-3, LNCaP, or DU-145 by small interfering RNA transfection caused significant reduction in proliferation and anchorage-independent growth on soft agar. This phenotype was associated with increased nuclear levels of the cyclin-dependent kinase inhibitor protein p27(Kip1) and diminished expression of S-phase promoting cyclin A2 and M-phase promoting cyclin B1 proteins. Finally, we show that elevated levels of FoxM1 protein correlate with high proliferation rates in human prostate adenocarcinomas. Our results suggest that the FoxM1 transcription factor regulates development and proliferation of prostate tumors, and that FoxM1 is a novel target for prostate cancer treatment.
Figures



References
-
- McCormick F. Signalling networks that cause cancer. Trends Cell Biol. 1999;9(12):M53–M6. - PubMed
-
- Franke TF, Hornik CP, Segev L, Shostak GA, Sugimoto C. PI3K/Akt and apoptosis: size matters. Oncogene. 2003;22(56):8983–98. - PubMed
-
- Sherr CJ, McCormick F. The RB and p53 pathways in cancer. Cancer Cell. 2002;2(2):103–12. - PubMed
-
- Massague J. G1 cell-cycle control and cancer. Nature. 2004;432(7015):298–306. - PubMed
-
- Sherr CJ, Roberts JM. CDK inhibitors: positive and negative regulators of G1-phase progression. Genes Dev. 1999;13(12):1501–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
