

Selective and antagonistic functions of SWI/SNF and Mi-2beta nucleosome remodeling complexes during an inflammatory response
- PMID: 16452502
- PMCID: PMC1361700
- DOI: 10.1101/gad.1383206
Selective and antagonistic functions of SWI/SNF and Mi-2beta nucleosome remodeling complexes during an inflammatory response
Abstract
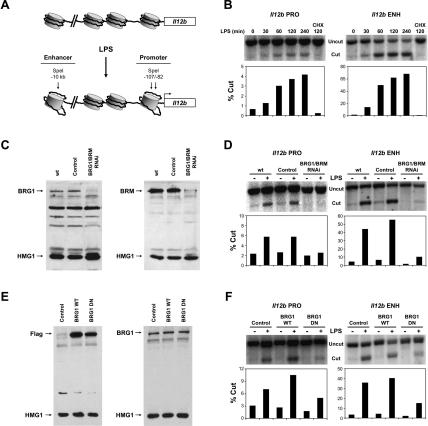
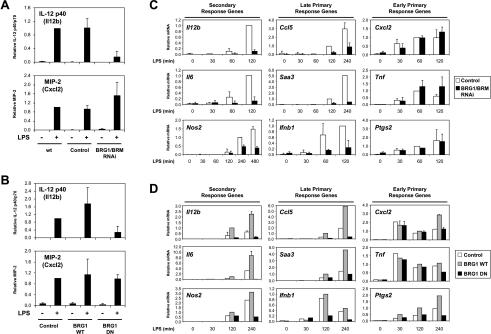
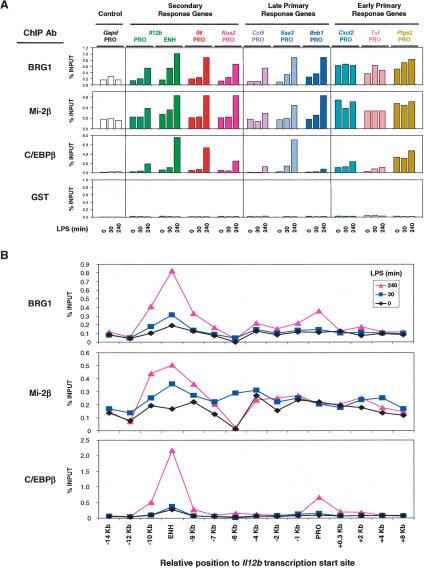
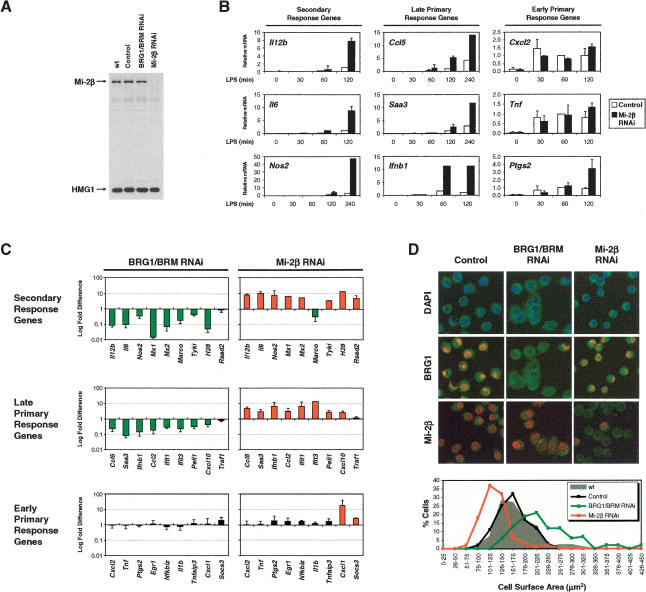
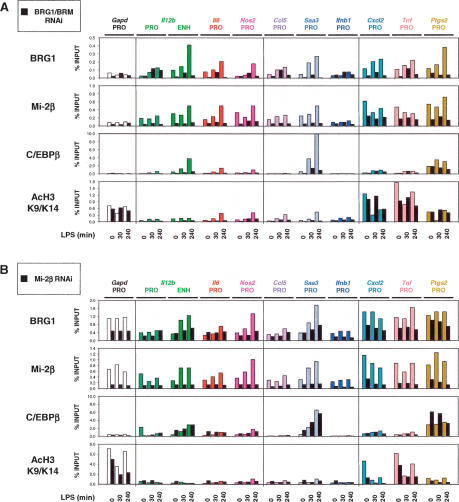
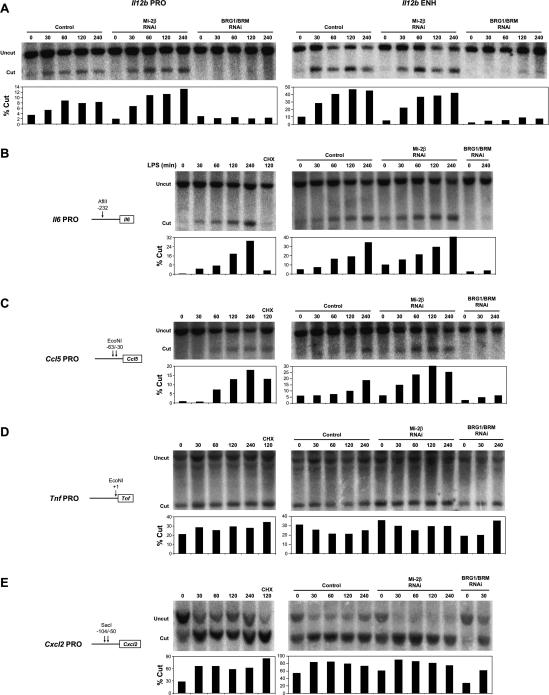
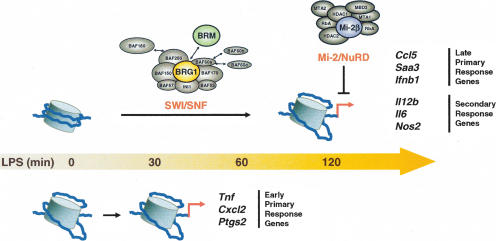
Studies of mammalian genes activated in response to an acute stimulus have suggested diverse mechanisms through which chromatin structure and nucleosome remodeling events contribute to inducible gene transcription. However, because of this diversity, the logical organization of the genome with respect to nucleosome remodeling and gene induction has remained obscure. Numerous proinflammatory genes are rapidly induced in macrophages in response to microbial infection. Here, we show that in lipopolysaccharide-stimulated macrophages, the catalytic BRG1/BRM subunits of the SWI/SNF class of ATP-dependent nucleosome remodeling complexes are consistently required for the activation of secondary response genes and primary response genes induced with delayed kinetics, but not for rapidly induced primary response genes. Surprisingly, a Mi-2beta complex was selectively recruited along with the SWI/SNF complexes to the control regions of secondary response and delayed primary response genes, with the Mi-2beta complex acting antagonistically to limit the induction of these gene classes. SWI/SNF and Mi-2beta complexes influenced cell size in a similarly antagonistic manner. These results provide insight into the differential contributions of nucleosome remodeling complexes to the rapid induction of defined classes of mammalian genes and reveal a robust anti-inflammatory function of Mi-2beta.
Figures
References
-
- Agalioti T., Lomvardas, S., Parekh, B., Yie, J., Maniatis, T., and Thanos, D. 2000. Ordered recruitment of chromatin modifying and general transcription factors to the IFN-β promoter. Cell 103: 667-678. - PubMed
-
- Akira S., Takeda, K., and Kaisho, T. 2001. Toll-like receptors: Critical proteins linking innate and acquired immunity. Nat. Immunol. 2: 675-680. - PubMed
-
- Becker P.B. and Horz, W. 2002. ATP-dependent nucleosome remodeling. Annu. Rev. Biochem. 71: 247-273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous