

Critical role of nitric oxide-cGMP cascade in the formation of cAMP-dependent long-term memory
- PMID: 16452652
- PMCID: PMC1360131
- DOI: 10.1101/lm.130506
Critical role of nitric oxide-cGMP cascade in the formation of cAMP-dependent long-term memory
Abstract
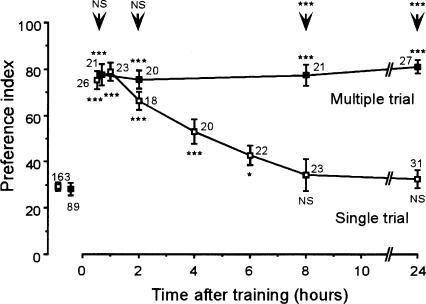
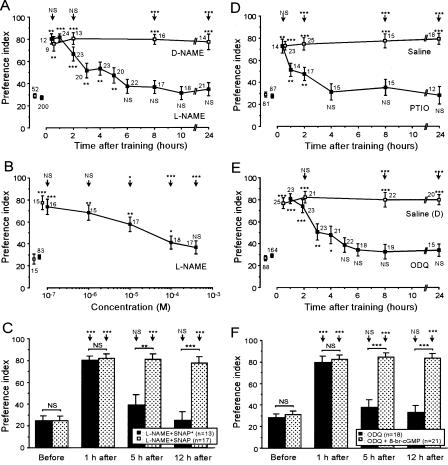
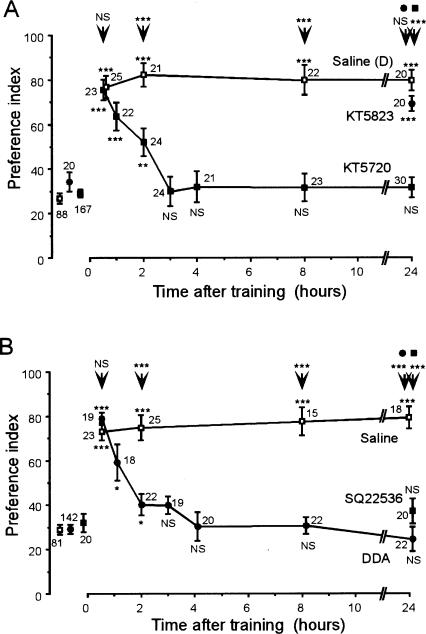
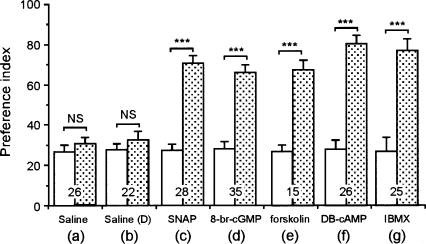
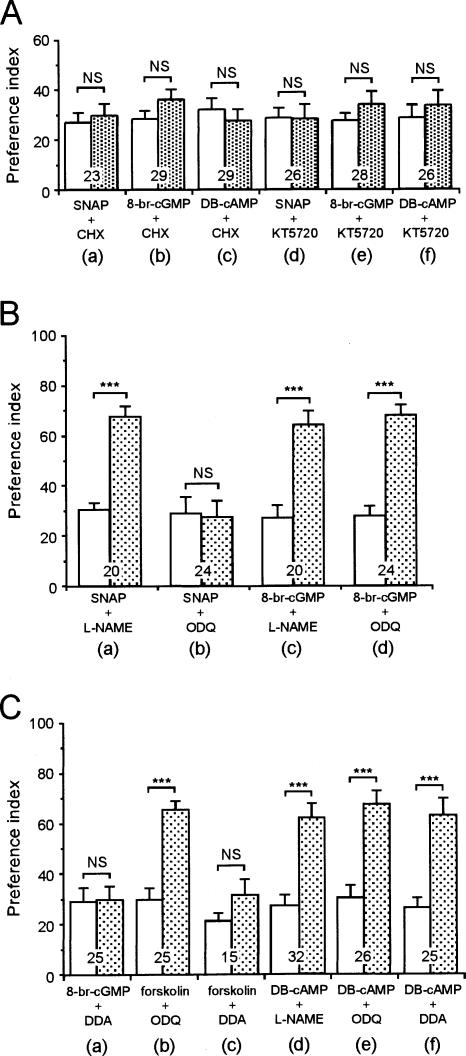
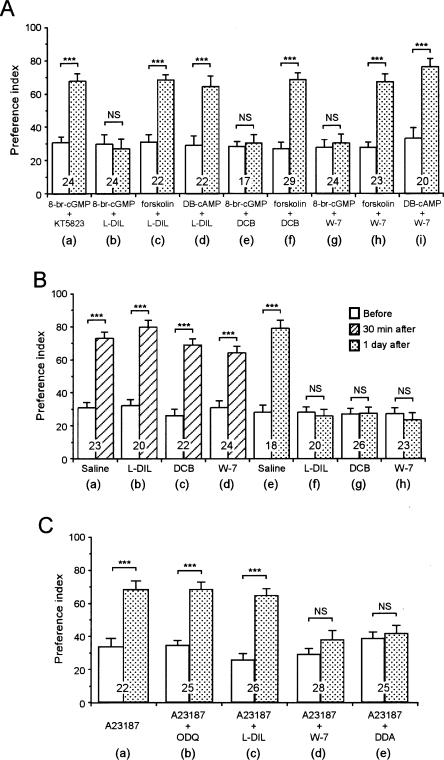
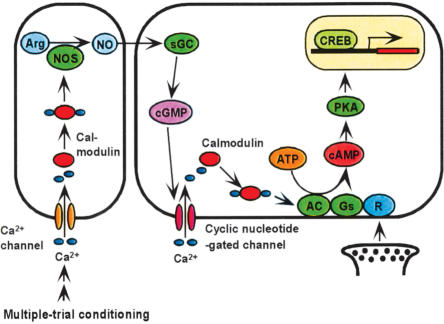
Cyclic AMP pathway plays an essential role in formation of long-term memory (LTM). In some species, the nitric oxide (NO)-cyclic GMP pathway has been found to act in parallel and complementary to the cAMP pathway for LTM formation. Here we describe a new role of the NO-cGMP pathway, namely, stimulation of the cAMP pathway to induce LTM. We have studied the signaling cascade underlying LTM formation by systematically coinjecting various "LTM-inducing" and "LTM-blocking" drugs in crickets. Multiple-trial olfactory conditioning led to LTM that lasted for several days, while memory induced by single-trial conditioning decayed away within several hours. Injection of inhibitors of the enzyme forming NO, cGMP, or cAMP into the hemolymph prior to multiple-trial conditioning blocked LTM, whereas injection of an NO donor, cGMP analog, or cAMP analog prior to single-trial conditioning induced LTM. Induction of LTM by injection of an NO donor or cGMP analog paired with single-trial conditioning was blocked by inhibitors of the cAMP pathway, but induction of LTM by a cAMP analog was unaffected by inhibitors of the NO-cGMP pathway. Inhibitors of cyclic nucleotide-gated channel (CNG channel) or calmodulin-blocked induction of LTM by cGMP analog paired with single-trial conditioning, but they did not affect induction of LTM by cAMP analog. Our findings suggest that the cAMP pathway is a downstream target of the NO-cGMP pathway for the formation of LTM, and that the CNG channel and calcium-calmodulin intervene between the NO-cGMP pathway and the cAMP pathway.
Figures
References
-
- Abel, T., Nguyen, P.V., Barad, M., Deuel, T.A.S., and Kandel, E.R. 1997. Genetic demonstration of a role for PKA in the late phase of LTP and in hippocampus-based long-term memory. Cell 88: 615-626. - PubMed
-
- Aonuma, H. and Newland, P.L. 2001. Opposing actions of nitric oxide on synaptic inputs of identified interneurones in the central nervous system of the crayfish. J. Exp. Biol. 204: 1319-1332. - PubMed
-
- ____. 2002. Synaptic inputs onto spiking local interneurons in crayfish are depressed by nitric oxide. J. Neurobiol. 52: 144-155. - PubMed
-
- Aonuma, H. and Niwa, K. 2004. Nitric oxide regulates the levels of cGMP accumulation in the cricket brain. Acta Biol. Hung. 55: 65-70. - PubMed
-
- Bartsch, D., Gjorardo, M., Skehel, P.A., Karl, K.A., Herder, S.P., Chen, M., Bailey, C.H., and Kandel, E.R. 1995. Aplysia CREB2 represses long-term facilitation: Relief of repression converts transient facilitation into long-term functional and structural change. Cell 83: 979-992. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical