

Physiological temperatures reduce the rate of vesicle pool depletion and short-term depression via an acceleration of vesicle recruitment
- PMID: 16452660
- PMCID: PMC6675486
- DOI: 10.1523/JNEUROSCI.3889-05.2006
Physiological temperatures reduce the rate of vesicle pool depletion and short-term depression via an acceleration of vesicle recruitment
Abstract
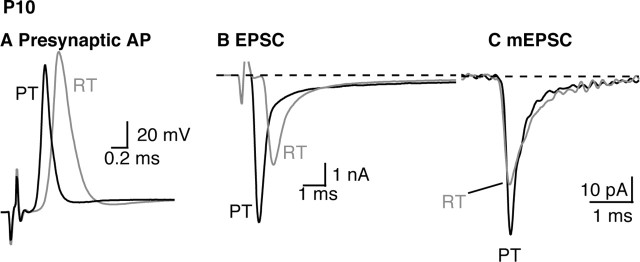
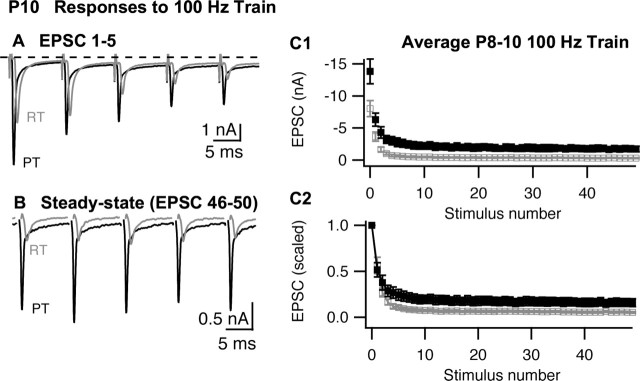
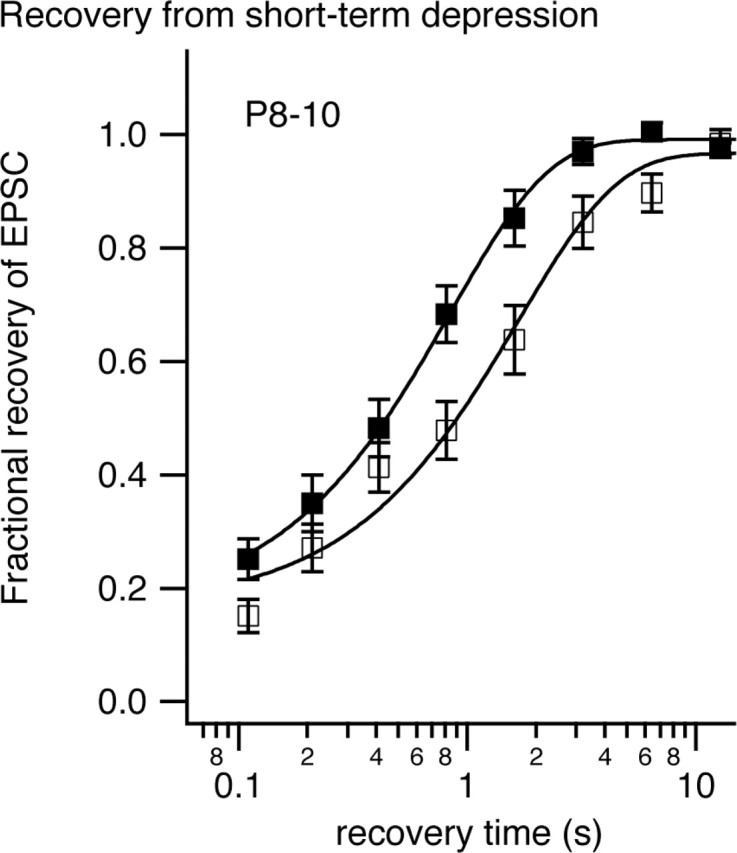
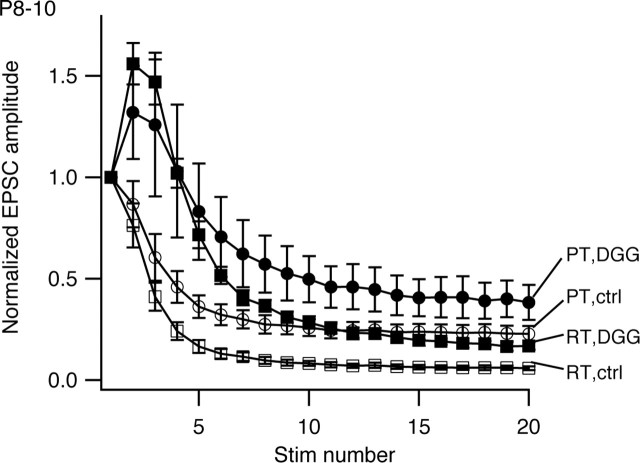
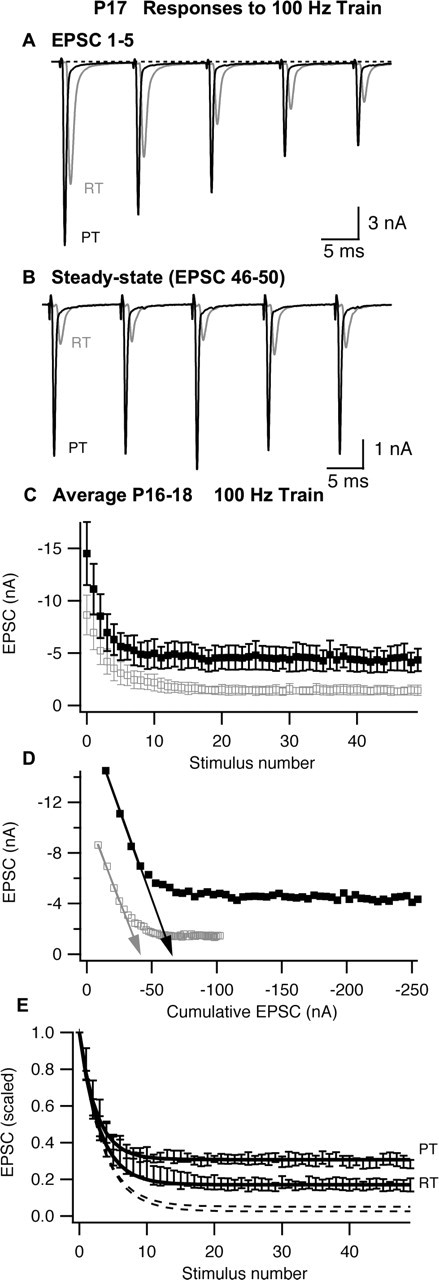
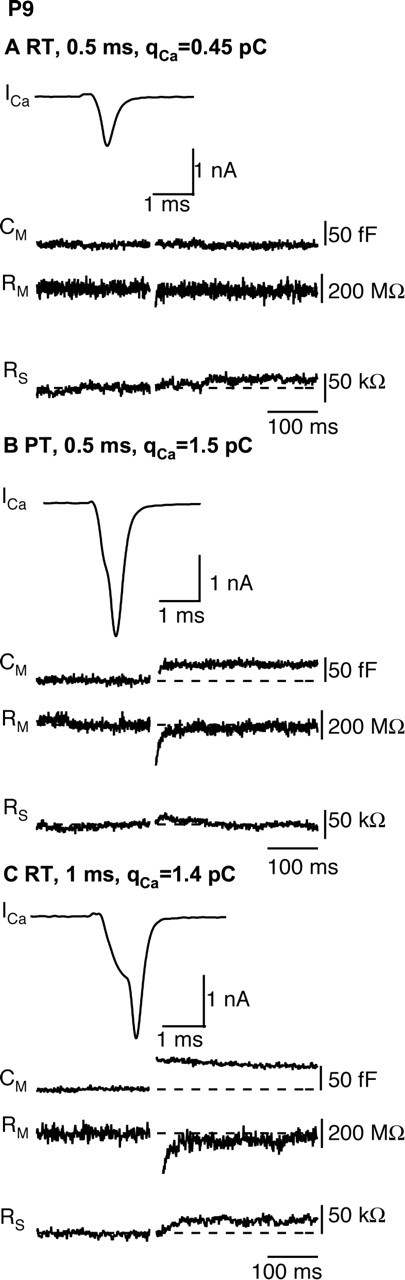
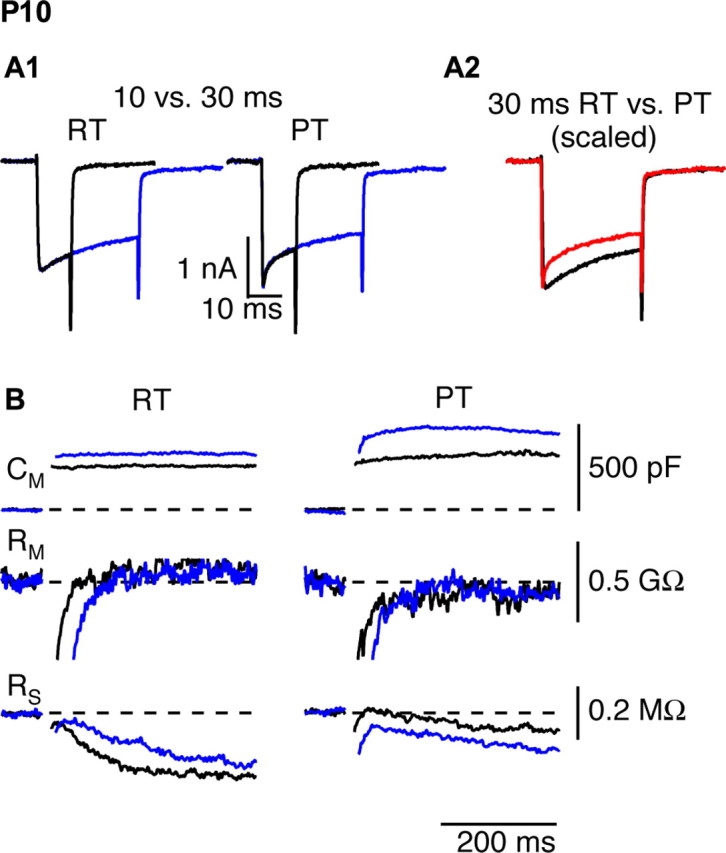
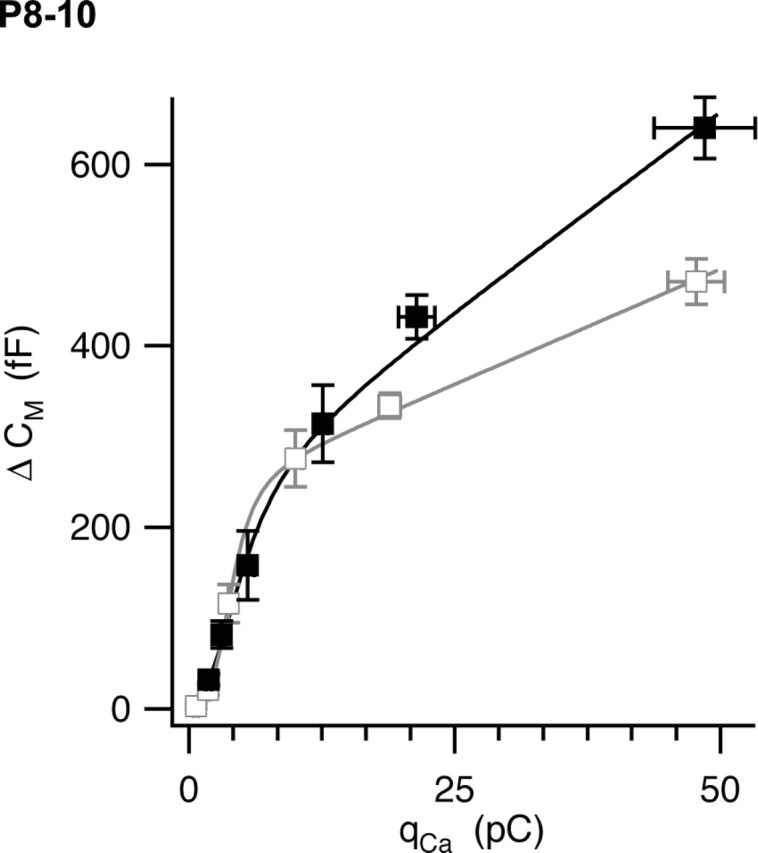
The timing and strength of synaptic transmission is profoundly dependent on temperature. However, the temperature dependence of the multiple mechanisms that contribute to short-term synaptic plasticity is poorly understood. Here, we use voltage-clamp recordings to quantify the temperature dependence of exocytosis at the calyx of Held synapse. EPSC and miniature EPSC amplitudes were larger at physiological temperature, but quantal content during low-frequency (0.05 Hz) stimulation was constant after temperature jumps from 22-24 degrees C to 35-37 degrees C. The initial degree of EPSC depression during 100 Hz stimuli trains was unchanged with temperature, as were estimates of release probability and vesicle pool size. In contrast, physiological temperatures dramatically relieved depression measured after 40 stimuli at 100 Hz by increasing twofold the rate of recovery from depression. Presynaptic calyx recordings revealed that physiological temperature increased capacitance jumps resulting from 0.5 and 1 ms depolarizations by increasing Ca2+ influx. When Ca2+ entry was equalized at the two temperatures, exocytosis exhibited little temperature dependence for brief depolarizations. However, in response to longer depolarizations, raising temperature increased a slow phase of exocytosis, without affecting overall Ca2+ entry or the size of the readily releasable pool of vesicles. Higher temperatures also increased the rate of presynaptic Ca2+ current inactivation; nevertheless, the degree of steady-state EPSC depression was greatly reduced. Our results thus suggest that changes in steady-state EPSCs during stimulus trains at physiological temperature reflect larger quantal amplitudes and faster refilling of synaptic vesicle pools, leading to reduced short-term depression during prolonged high-frequency firing.
Figures
References
-
- Bollmann JH, Sakmann B, Borst JG (2000). Calcium sensitivity of glutamate release in a calyx-type terminal. Science 289:953–957. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous