

Pre-steady-state binding of damaged DNA by XPC-hHR23B reveals a kinetic mechanism for damage discrimination
- PMID: 16460043
- PMCID: PMC2435173
- DOI: 10.1021/bi051936t
Pre-steady-state binding of damaged DNA by XPC-hHR23B reveals a kinetic mechanism for damage discrimination
Abstract
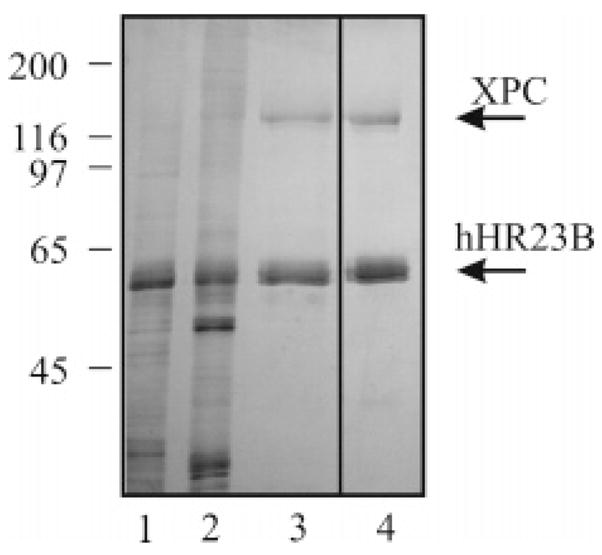
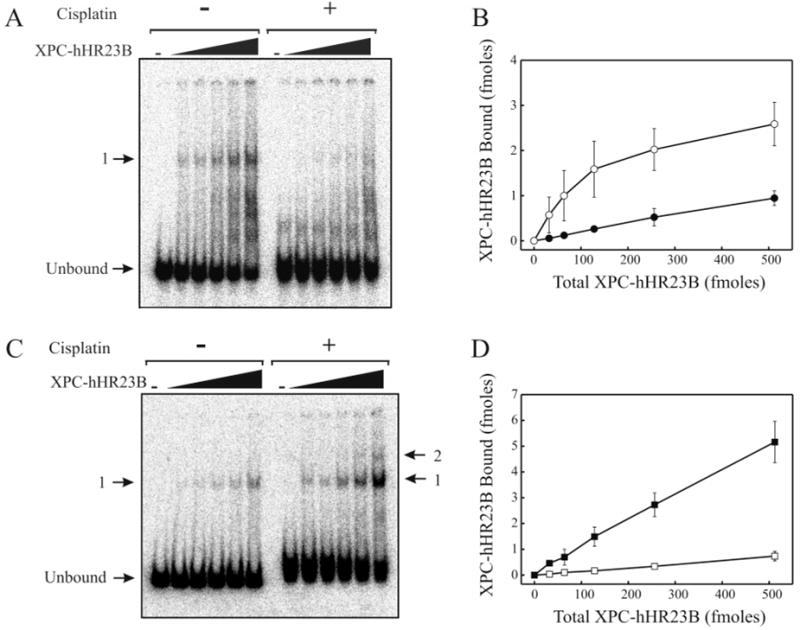
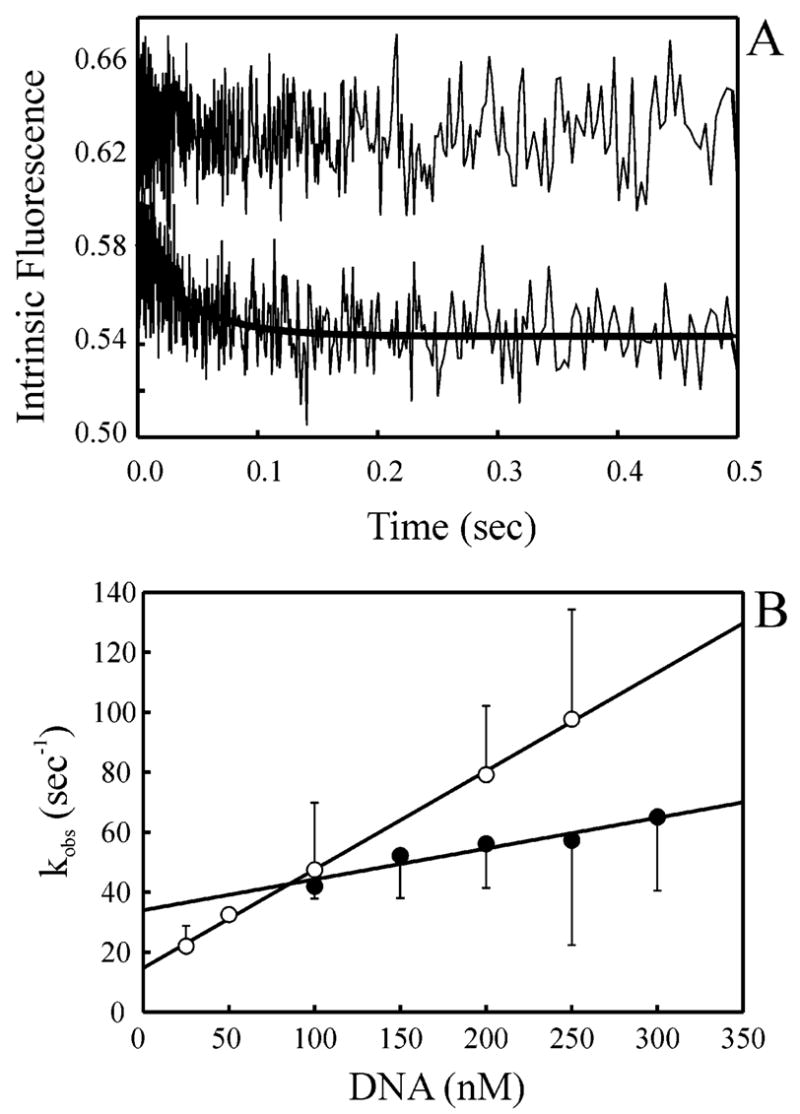
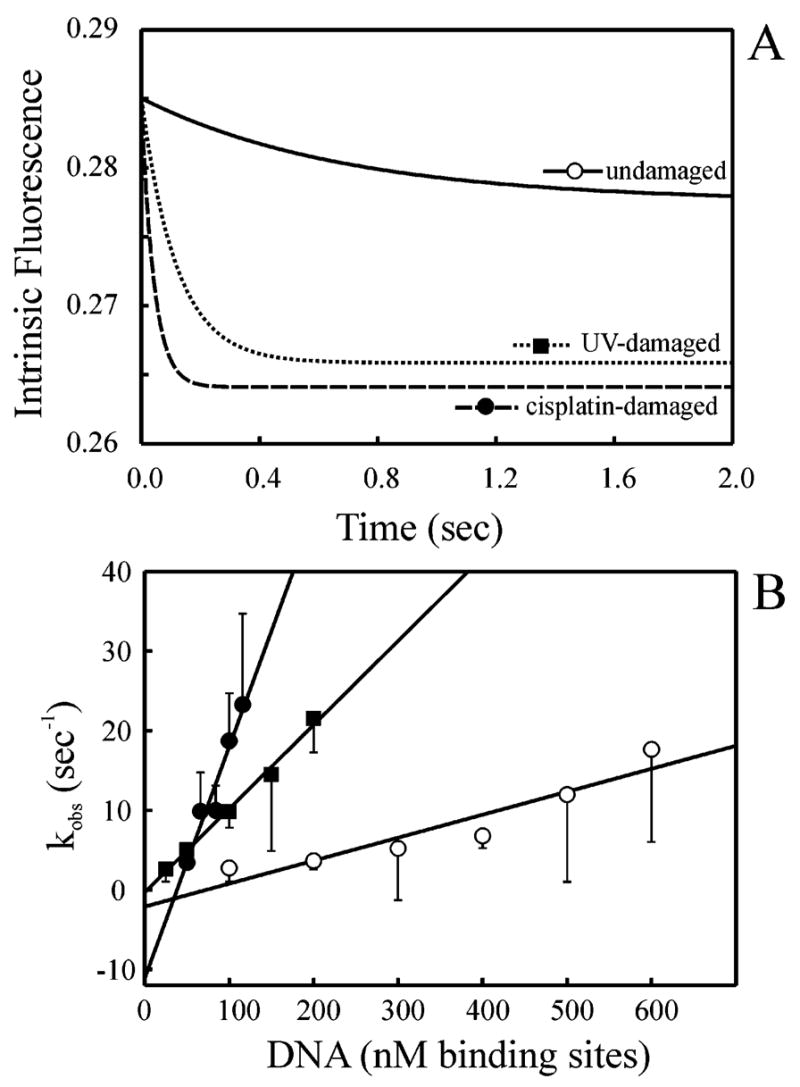
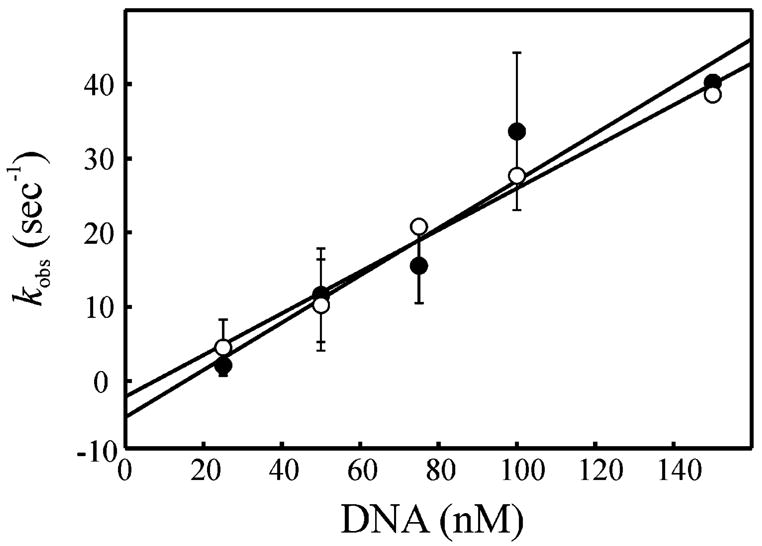
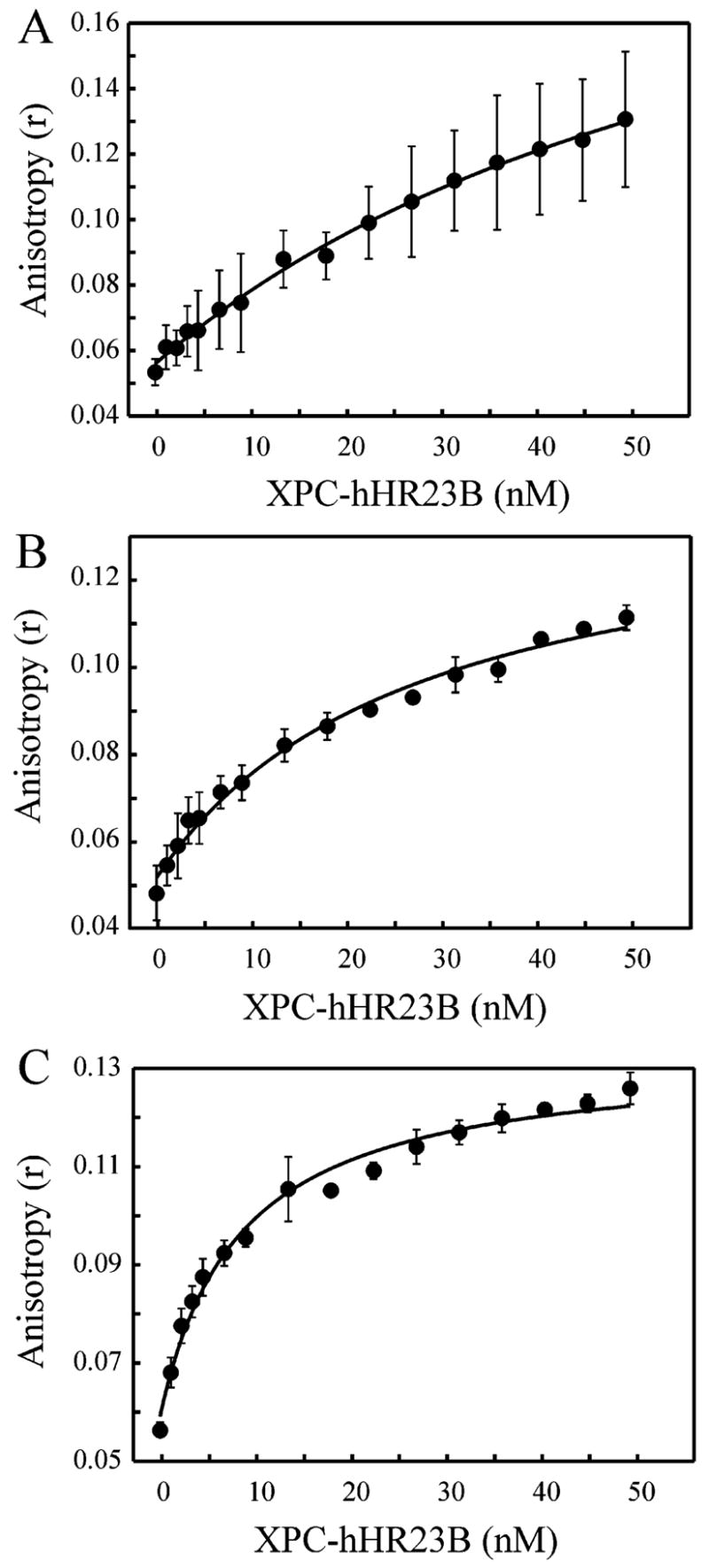
The XPC-hHR23B complex (XPC-hHR23B) is a heterodimeric protein required for the initial step of DNA damage recognition in the global nucleotide excision repair (NER) pathway. A strong preference of XPC-hHR23B for UV- and cisplatin-damaged DNA has previously been demonstrated using equilibrium binding assays. To better understand the molecular mechanism of damage recognition by XPC-hHR23B, we carried out the pre-steady-state kinetic analysis of the XPC-hHR23B-DNA interactions using a stopped-flow fluorescence assay. XPC-hHR23B displays a faster k(on) for cisplatin- and UV-damaged duplex DNA than for undamaged DNA, with additional, minor effects on the k(off) rates. XPC-hHR23B has a high affinity for undamaged single-stranded DNA compared to duplex DNA, which can be largely attributed to a high rate of association. However, cisplatin damage on single-stranded DNA reduced the overall level of binding by a factor of 7, with nearly equal contributions from changes to the k(on) and k(off) rates. Together, these results support a model for initial damage recognition by XPC-hHR23B that is dependent on structural changes in the DNA, and not adduct chemistry.
Figures
Similar articles
-
Stable binding of human XPC complex to irradiated DNA confers strong discrimination for damaged sites.J Mol Biol. 2000 Jul 7;300(2):275-90. doi: 10.1006/jmbi.2000.3857. J Mol Biol. 2000. PMID: 10873465
-
Biochemical analysis of the damage recognition process in nucleotide excision repair.J Biol Chem. 2003 Feb 28;278(9):7476-85. doi: 10.1074/jbc.M210603200. Epub 2002 Dec 13. J Biol Chem. 2003. PMID: 12486030
-
Human XPC-hHR23B interacts with XPA-RPA in the recognition of triplex-directed psoralen DNA interstrand crosslinks.Nucleic Acids Res. 2005 May 24;33(9):2993-3001. doi: 10.1093/nar/gki610. Print 2005. Nucleic Acids Res. 2005. PMID: 15914671 Free PMC article.
-
Critical DNA damage recognition functions of XPC-hHR23B and XPA-RPA in nucleotide excision repair.Mol Carcinog. 2003 Sep;38(1):1-13. doi: 10.1002/mc.10143. Mol Carcinog. 2003. PMID: 12949838 Review.
-
DNA damage recognition during nucleotide excision repair in mammalian cells.Biochimie. 1999 Jan-Feb;81(1-2):39-44. doi: 10.1016/s0300-9084(99)80036-4. Biochimie. 1999. PMID: 10214908 Review.
Cited by
-
Enhanced spontaneous DNA twisting/bending fluctuations unveiled by fluorescence lifetime distributions promote mismatch recognition by the Rad4 nucleotide excision repair complex.Nucleic Acids Res. 2018 Feb 16;46(3):1240-1255. doi: 10.1093/nar/gkx1216. Nucleic Acids Res. 2018. PMID: 29267981 Free PMC article.
-
Polynuclear ruthenium organometallic compounds induce DNA damage in human cells identified by the nucleotide excision repair factor XPC.Biosci Rep. 2019 Jul 15;39(7):BSR20190378. doi: 10.1042/BSR20190378. Print 2019 Jul 31. Biosci Rep. 2019. PMID: 31227614 Free PMC article.
-
Two-stage dynamic DNA quality check by xeroderma pigmentosum group C protein.EMBO J. 2009 Aug 19;28(16):2387-99. doi: 10.1038/emboj.2009.187. Epub 2009 Jul 16. EMBO J. 2009. PMID: 19609301 Free PMC article.
-
Twist-open mechanism of DNA damage recognition by the Rad4/XPC nucleotide excision repair complex.Proc Natl Acad Sci U S A. 2016 Apr 19;113(16):E2296-305. doi: 10.1073/pnas.1514666113. Epub 2016 Mar 31. Proc Natl Acad Sci U S A. 2016. PMID: 27035942 Free PMC article.
-
Strand- and site-specific DNA lesion demarcation by the xeroderma pigmentosum group D helicase.Proc Natl Acad Sci U S A. 2010 Oct 12;107(41):17545-50. doi: 10.1073/pnas.1004339107. Epub 2010 Sep 27. Proc Natl Acad Sci U S A. 2010. PMID: 20876134 Free PMC article.
References
-
- Aboussekhra A, Biggerstaff M, Shivji MK, Vilpo JA, Moncollin V, Podust VN, Protic M, Hubscher U, Egly JM, Wood RD. Mammalian DNA nucleotide excision repair reconstituted with purified protein components. Cell. 1995;80:859–868. - PubMed
-
- Mu D, Park CH, Matsunaga T, Hsu DS, Reardon JT, Sancar A. Reconstitution of human DNA repair excision nuclease in a highly defined system. J Biol Chem. 1995;270:2415–2418. - PubMed
-
- Hanawalt PC. DNA damage and repair. Volume 2: DNA repair in higher eukaryotes Overview. DNA Damage Repair. 1998;2:1–8.
-
- Batty DP, Wood RD. Damage recognition in nucleotide excision repair of DNA. Gene. 2000;241:193–204. - PubMed
-
- Christians FC, Hanawalt PC. Repair in ribosomal-RNA genes is deficient in xeroderma pigmentosum group-C and in cockaynes-syndrome cells. Mutat Res. 1994;323:179–187. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
