

Brainstem circuits regulating gastric function
- PMID: 16460274
- PMCID: PMC3062484
- DOI: 10.1146/annurev.physiol.68.040504.094635
Brainstem circuits regulating gastric function
Abstract
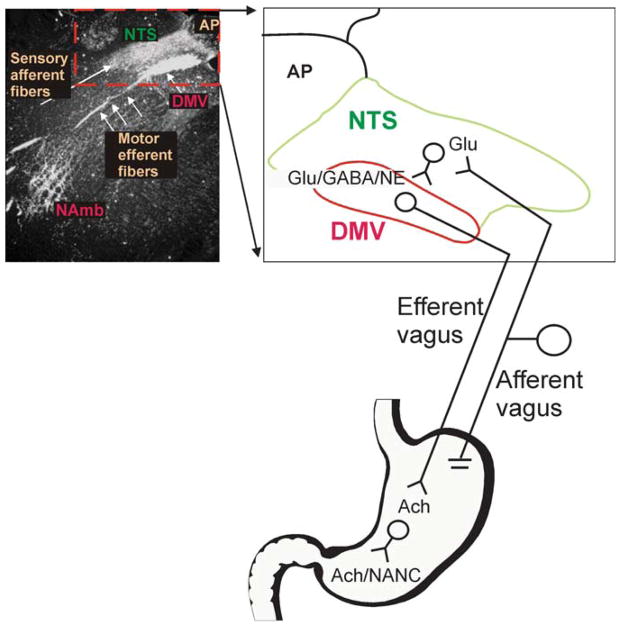
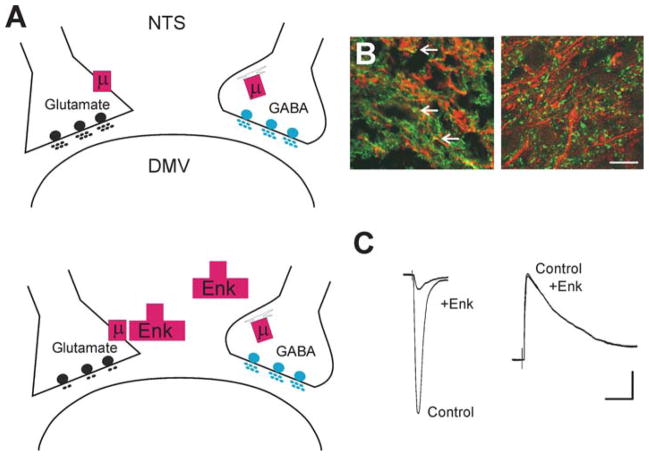
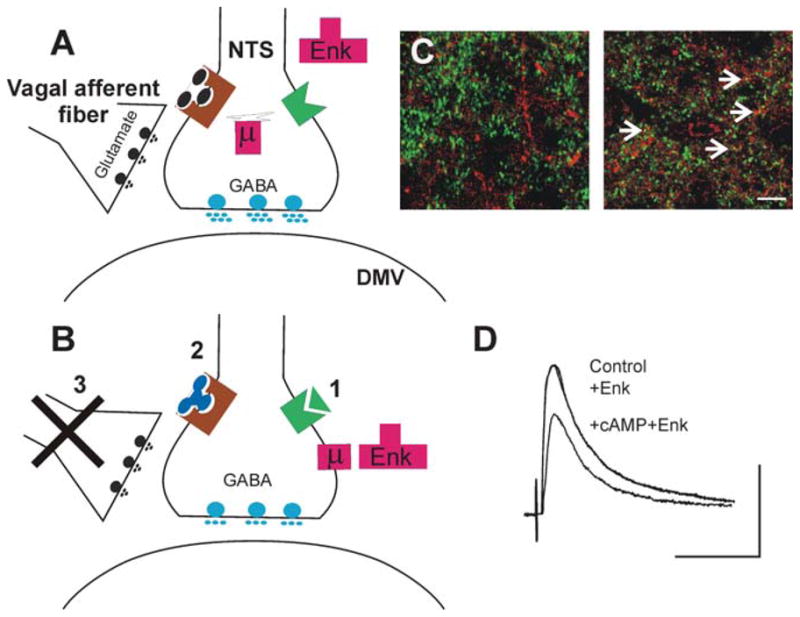
Brainstem parasympathetic circuits that modulate digestive functions of the stomach are comprised of afferent vagal fibers, neurons of the nucleus tractus solitarius (NTS), and the efferent fibers originating in the dorsal motor nucleus of the vagus (DMV). A large body of evidence has shown that neuronal communications between the NTS and the DMV are plastic and are regulated by the presence of a variety of neurotransmitters and circulating hormones as well as the presence, or absence, of afferent input to the NTS. These data suggest that descending central nervous system inputs as well as hormonal and afferent feedback resulting from the digestive process can powerfully regulate vago-vagal reflex sensitivity. This paper first reviews the essential "static" organization and function of vago-vagal gastric control neurocircuitry. We then present data on the opioidergic modulation of NTS connections with the DMV as an example of the "gating" of these reflexes, i.e., how neurotransmitters, hormones, and vagal afferent traffic can make an otherwise static autonomic reflex highly plastic.
Figures
References
-
- Wood JD. Physiology of the enteric nervous system. In: Johnson LR, editor. Physiology of the Gastrointestinal Tract. New York: Raven; 1987. pp. 67–110.
-
- Goyal RK, Hirano I. The enteric nervous system. N Engl J Med. 1996;334:1106–15. - PubMed
-
- Bornstein JC, Costa M, Grider JR. Enteric motor and interneuronal circuits controlling motility. Neurogastroenterol Motil. 2004;16(Suppl 1):34–38. - PubMed
-
- Goyal RK, Paterson WG. Esophageal motility. In: Wood JD, editor. Handbook of Physiology—The Gastrointestinal System I. Bethesda, MD: Am. Phys. Soc; 1989. pp. 865–908.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
