

Interaction of beta-1,3-glucan with its recognition protein activates hemolymph proteinase 14, an initiation enzyme of the prophenoloxidase activation system in Manduca sexta
- PMID: 16461344
- PMCID: PMC2020818
- DOI: 10.1074/jbc.M513797200
Interaction of beta-1,3-glucan with its recognition protein activates hemolymph proteinase 14, an initiation enzyme of the prophenoloxidase activation system in Manduca sexta
Abstract
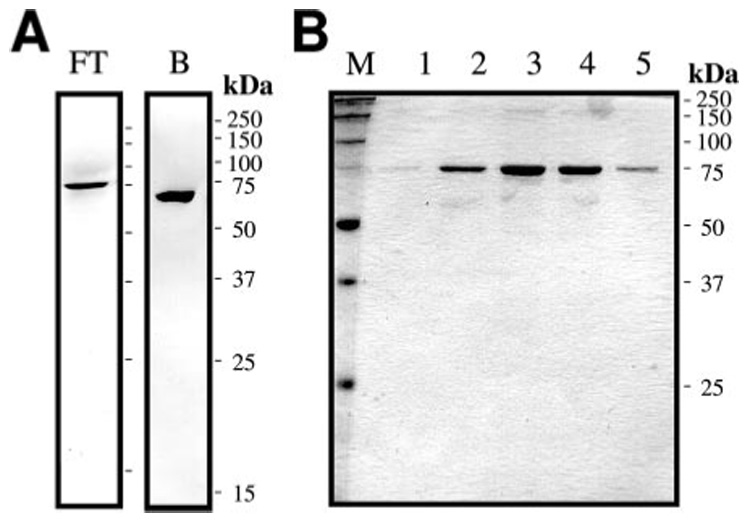
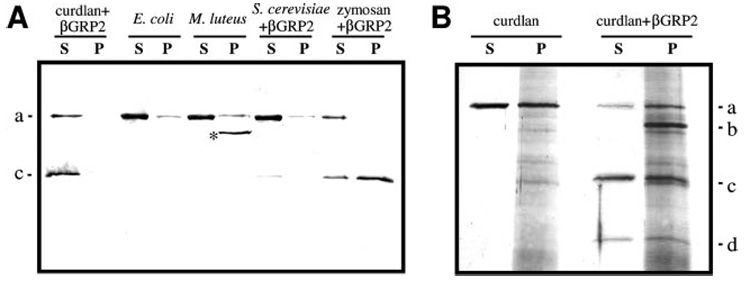
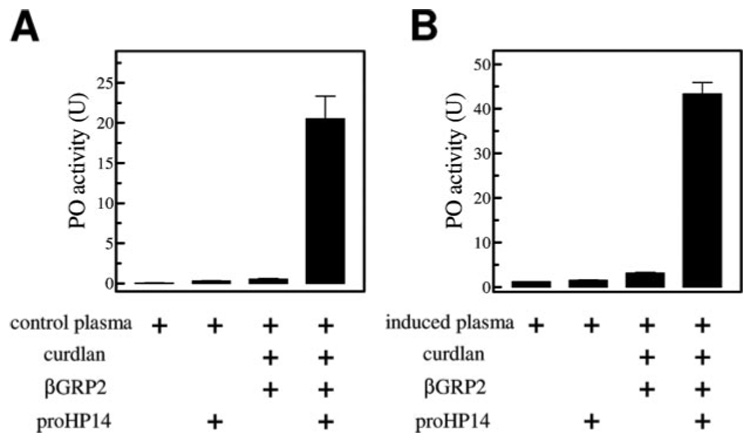
A serine proteinase pathway in insect hemolymph leads to prophenoloxidase activation, an innate immune response against pathogen infection. In the tobacco hornworm Manduca sexta, recombinant hemolymph proteinase 14 precursor (pro-HP14) interacts with peptidoglycan, autoactivates, and initiates the proteinase cascade (Ji, C., Wang, Y., Guo, X., Hartson, S., and Jiang, H. (2004) J. Biol. Chem. 279, 34101-34106). Here, we report the purification and characterization of pro-HP14 from the hemolymph of bacteria-injected M. sexta larvae. The zymogen, consisting of a single polypeptide with a molecular mass of 68.5 kDa, is truncated at the amino terminus. It is converted to a two-chain active form in the presence of beta-1,3-glucan (a fungal cell wall component) and beta-1,3-glucan recognition protein-2. The 45-kDa heavy chain contains four low-density lipoprotein receptor A repeats, one Sushi domain, and one unique cysteine-rich region, whereas the 30-kDa light chain contains a serine proteinase domain, which was labeled by [(3)H]diisopropyl fluorophosphate. Pro-HP14 in the plasma strongly binds curdlan, zymosan, and yeast and interacts with peptidoglycan and Micrococcus luteus. Addition of autoactivated HP14 elevated phenoloxidase activity level in the larval plasma. Recombinant M. sexta serpin-1I reduced prophenoloxidase activation by inhibiting HP14. These data are consistent with the current model on initiation and regulation of the prophenoloxidase activation cascade upon recognition of pathogen-associated molecular patterns by specific pattern recognition proteins.
Figures




Similar articles
-
Inhibition of immune pathway-initiating hemolymph protease-14 by Manduca sexta serpin-12, a conserved mechanism for the regulation of melanization and Toll activation in insects.Insect Biochem Mol Biol. 2020 Jan;116:103261. doi: 10.1016/j.ibmb.2019.103261. Epub 2019 Nov 4. Insect Biochem Mol Biol. 2020. PMID: 31698082 Free PMC article.
-
Reconstitution of a branch of the Manduca sexta prophenoloxidase activation cascade in vitro: snake-like hemolymph proteinase 21 (HP21) cleaved by HP14 activates prophenoloxidase-activating proteinase-2 precursor.Insect Biochem Mol Biol. 2007 Oct;37(10):1015-25. doi: 10.1016/j.ibmb.2007.05.013. Epub 2007 May 29. Insect Biochem Mol Biol. 2007. PMID: 17785189 Free PMC article.
-
A pattern recognition serine proteinase triggers the prophenoloxidase activation cascade in the tobacco hornworm, Manduca sexta.J Biol Chem. 2004 Aug 13;279(33):34101-6. doi: 10.1074/jbc.M404584200. Epub 2004 Jun 9. J Biol Chem. 2004. PMID: 15190055
-
Innate immune responses of a lepidopteran insect, Manduca sexta.Immunol Rev. 2004 Apr;198:97-105. doi: 10.1111/j.0105-2896.2004.0121.x. Immunol Rev. 2004. PMID: 15199957 Review.
-
Pattern recognition proteins in Manduca sexta plasma.Insect Biochem Mol Biol. 2002 Oct;32(10):1287-93. doi: 10.1016/s0965-1748(02)00091-7. Insect Biochem Mol Biol. 2002. PMID: 12225919 Review.
Cited by
-
Inhibition of immune pathway-initiating hemolymph protease-14 by Manduca sexta serpin-12, a conserved mechanism for the regulation of melanization and Toll activation in insects.Insect Biochem Mol Biol. 2020 Jan;116:103261. doi: 10.1016/j.ibmb.2019.103261. Epub 2019 Nov 4. Insect Biochem Mol Biol. 2020. PMID: 31698082 Free PMC article.
-
Immunity in lepidopteran insects.Adv Exp Med Biol. 2010;708:181-204. doi: 10.1007/978-1-4419-8059-5_10. Adv Exp Med Biol. 2010. PMID: 21528699 Free PMC article. Review.
-
Insect prophenoloxidase: the view beyond immunity.Front Physiol. 2014 Jul 11;5:252. doi: 10.3389/fphys.2014.00252. eCollection 2014. Front Physiol. 2014. PMID: 25071597 Free PMC article. Review.
-
Initiating protease with modular domains interacts with β-glucan recognition protein to trigger innate immune response in insects.Proc Natl Acad Sci U S A. 2015 Nov 10;112(45):13856-61. doi: 10.1073/pnas.1517236112. Epub 2015 Oct 26. Proc Natl Acad Sci U S A. 2015. PMID: 26504233 Free PMC article.
-
Hemolymph protease-5 links the melanization and Toll immune pathways in the tobacco hornworm, Manduca sexta.Proc Natl Acad Sci U S A. 2020 Sep 22;117(38):23581-23587. doi: 10.1073/pnas.2004761117. Epub 2020 Sep 8. Proc Natl Acad Sci U S A. 2020. PMID: 32900946 Free PMC article.
References
-
- Gillespie JP, Kanost MR, Trenczek T. Annu. Rev. Entomol. 1997;42:611–643. - PubMed
-
- Hoffmann JA. Nature. 2003;426:33–38. - PubMed
-
- Lavine MD, Strand MR. Insect Biochem. Mol. Biol. 2002;32:1295–1309. - PubMed
-
- Jiang H, Kanost MR. Insect Biochem. Mol. Biol. 2000;30:95–105. - PubMed
-
- Iwanaga S, Kawabata S, Muta T. J. Biochem. 1998;123:1–15. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
