

Subtelomeric proteins negatively regulate telomere elongation in budding yeast
- PMID: 16467854
- PMCID: PMC1383556
- DOI: 10.1038/sj.emboj.7600975
Subtelomeric proteins negatively regulate telomere elongation in budding yeast
Abstract
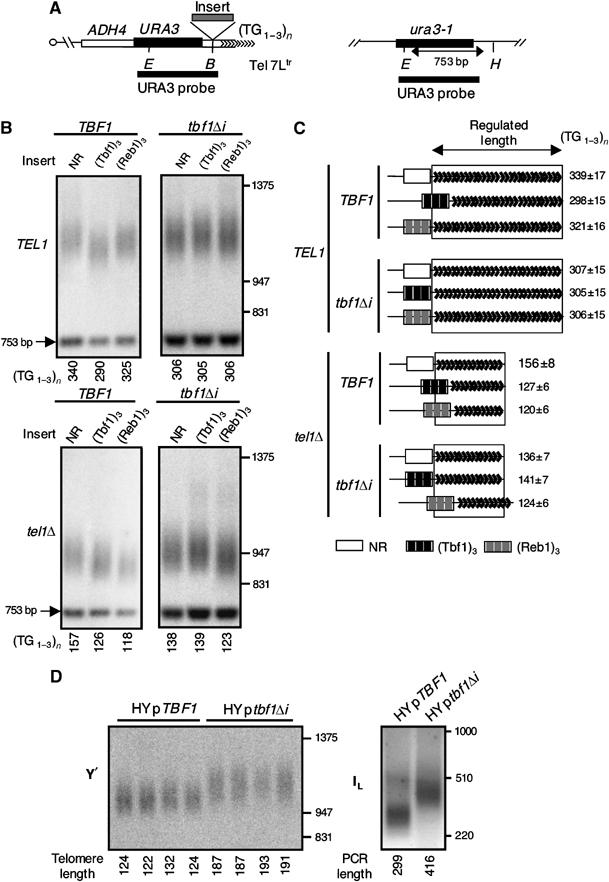
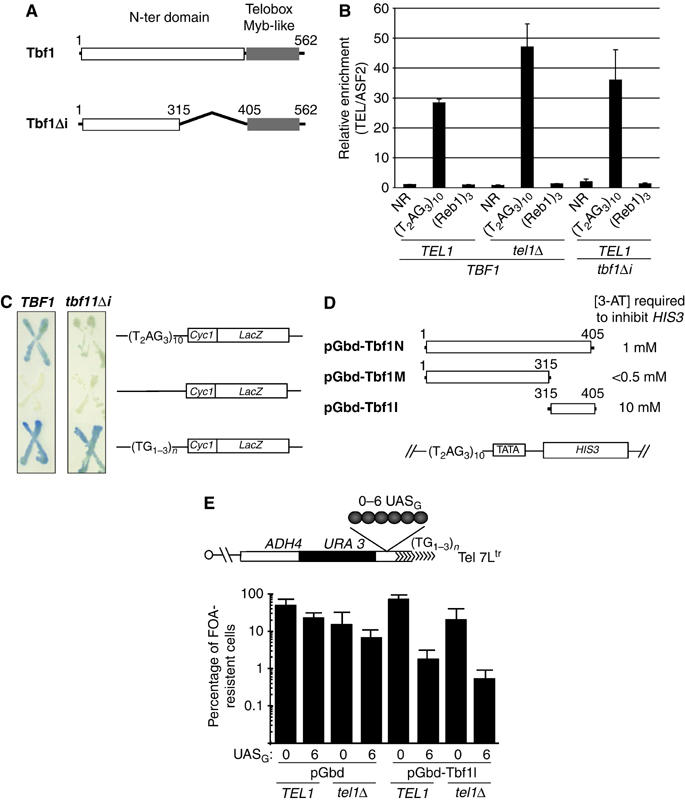
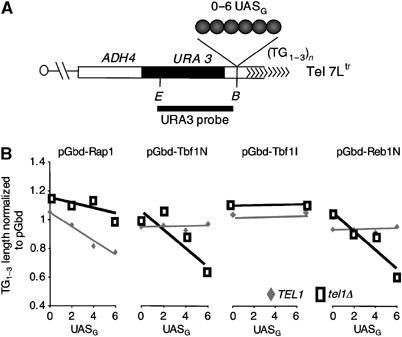
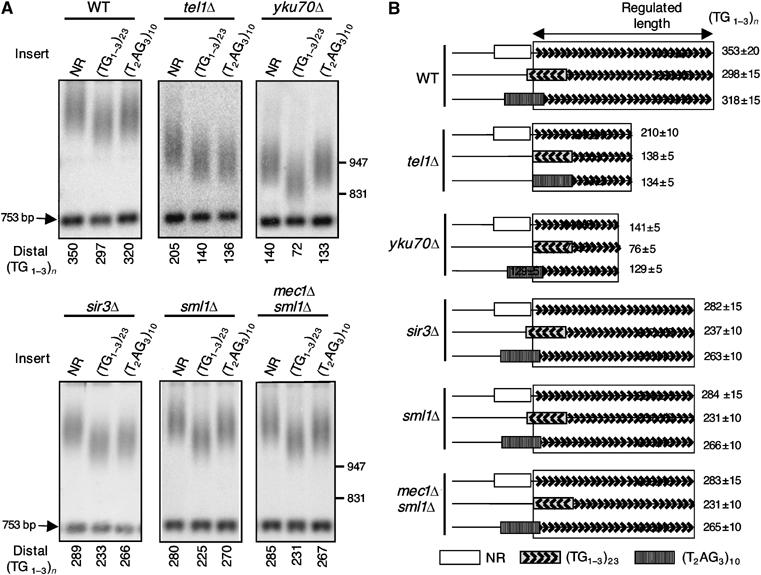
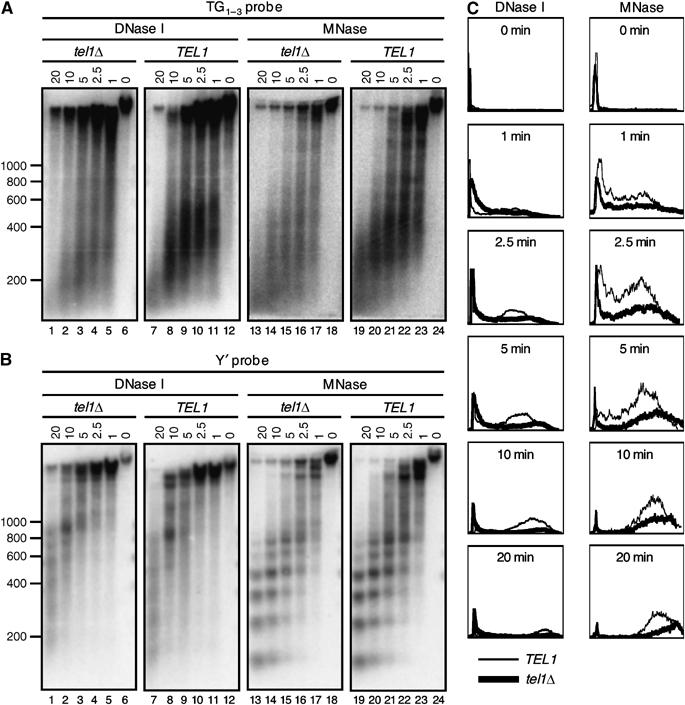
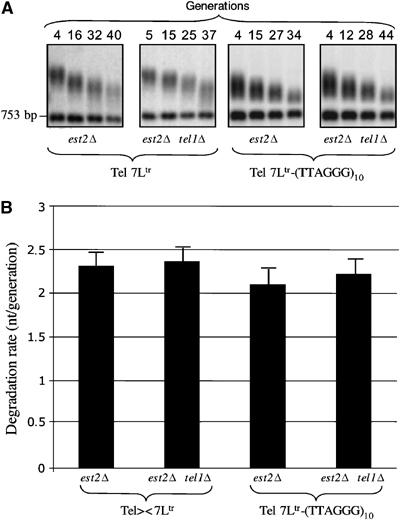
The Tbf1 and Reb1 proteins are present in yeast subtelomeric regions. We establish in this work that they inhibit telomerase-dependent lengthening of telomere. For example, tethering the N-terminal domain of Tbf1 and Reb1 in a subtelomeric region shortens that telomere proportionally to the number of domains bound. We further identified a 90 amino-acid long sequence within the N-terminal domain of Tbf1 that is necessary but not sufficient for its length regulation properties. The role of the subtelomeric factors in telomere length regulation is antagonized by TEL1 and does not correlate with a global telomere derepression. We show that the absence of TEL1 induces an alteration in the structure of telomeric chromatin, as defined biochemically by an increased susceptibility to nucleases and a greater heterogeneity of products. We propose that the absence of TEL1 modifies the organization of the telomeres, which allows Tbf1 and Reb1 to cis-inhibit telomerase. The involvement of subtelomeric factors in telomere length regulation provides a possible mechanism for the chromosome-specific length setting observed at yeast and human telomeres.
Figures
References
-
- Ancelin K, Brunori M, Bauwens S, Koering CE, Brun C, Ricoul M, Pommier JP, Sabatier L, Gilson E (2002) Targeting assay to study the cis functions of human telomeric proteins: evidence for inhibition of telomerase by TRF1 and for activation of telomere degradation by TRF2. Mol Cell Biol 22: 3474–3487 - PMC - PubMed
-
- Aparicio OM, Billington BL, Gottschling DE (1991) Modifiers of position effect are shared between telomeric and silent mating-type loci in S. cerevisiae. Cell 66: 1279–1287 - PubMed
-
- Blackburn EH (1991) Structure and function of telomeres. Nature 18: 569–573 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
