

FLOWERING LOCUS C mediates natural variation in the high-temperature response of the Arabidopsis circadian clock
- PMID: 16473970
- PMCID: PMC1383639
- DOI: 10.1105/tpc.105.038315
FLOWERING LOCUS C mediates natural variation in the high-temperature response of the Arabidopsis circadian clock
Abstract
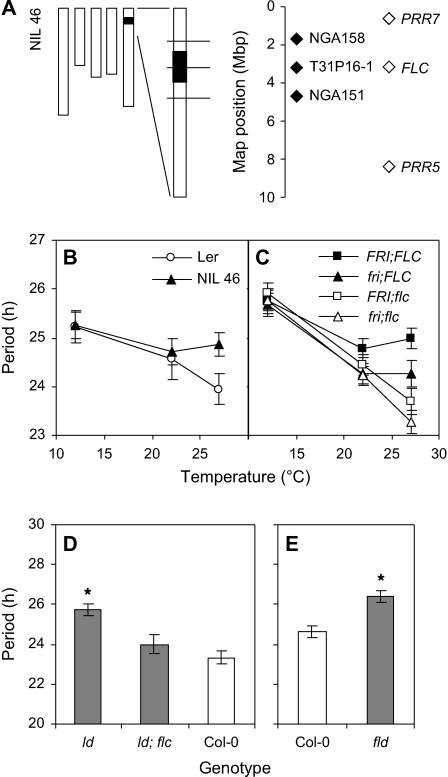
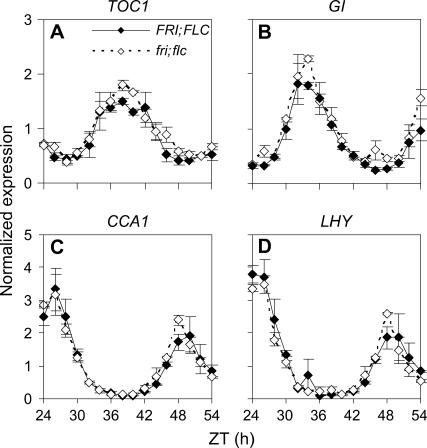
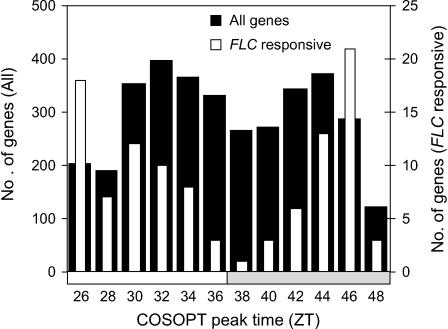
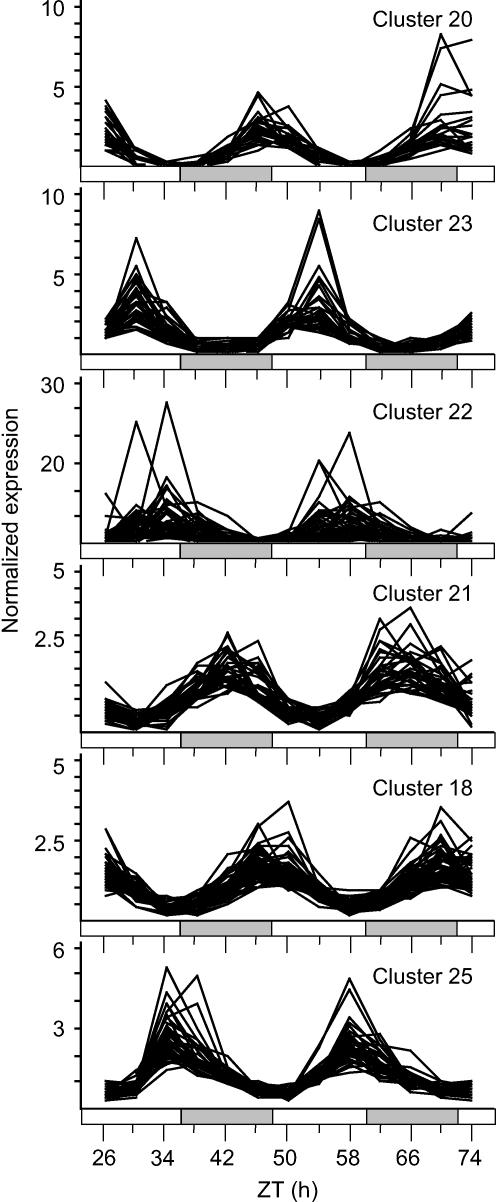
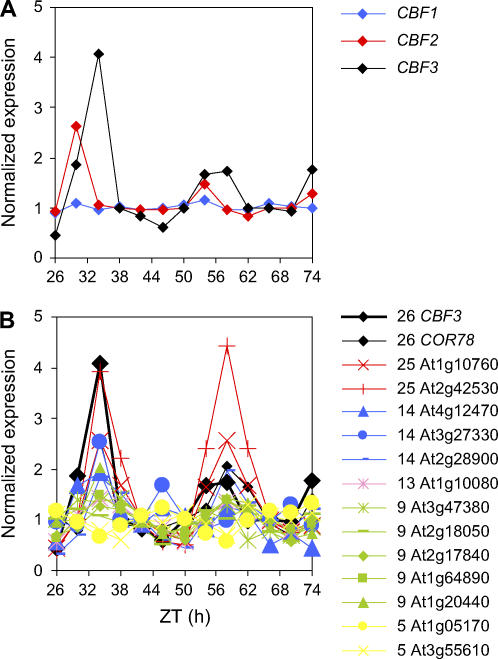
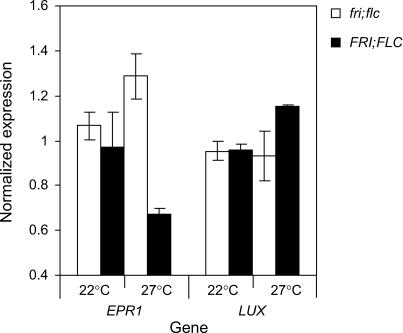
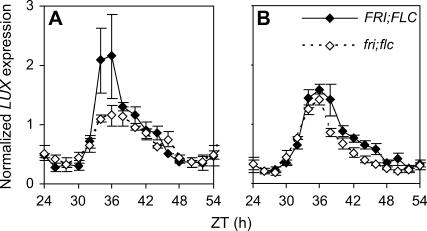
Temperature compensation contributes to the accuracy of biological timing by preventing circadian rhythms from running more quickly at high than at low temperatures. We previously identified quantitative trait loci (QTL) with temperature-specific effects on the circadian rhythm of leaf movement, including a QTL linked to the transcription factor FLOWERING LOCUS C (FLC). We have now analyzed FLC alleles in near-isogenic lines and induced mutants to eliminate other candidate genes. We showed that FLC lengthened the circadian period specifically at 27 degrees C, contributing to temperature compensation of the circadian clock. Known upstream regulators of FLC expression in flowering time pathways similarly controlled its circadian effect. We sought to identify downstream targets of FLC regulation in the molecular mechanism of the circadian clock using genome-wide analysis to identify FLC-responsive genes and 3503 transcripts controlled by the circadian clock. A Bayesian clustering method based on Fourier coefficients allowed us to discriminate putative regulatory genes. Among rhythmic FLC-responsive genes, transcripts of the transcription factor LUX ARRHYTHMO (LUX) correlated in peak abundance with the circadian period in flc mutants. Mathematical modeling indicated that the modest change in peak LUX RNA abundance was sufficient to cause the period change due to FLC, providing a molecular target for the crosstalk between flowering time pathways and circadian regulation.
Figures
References
-
- Akhtar, R.A., Reddy, A.B., Maywood, E.S., Clayton, J.D., King, V.M., Smith, A.G., Gant, T.W., Hastings, M.H., and Kyriacou, C.P. (2002). Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr. Biol. 12 540–550. - PubMed
-
- Alabadi, D., Oyama, T., Yanovsky, M.J., Harmon, F.G., Mas, P., and Kay, S.A. (2001). Reciprocal regulation between TOC1 and LHY/CCA1. Science 293 880–883. - PubMed
-
- Anderson, P.E., Smith, J.Q., Edwards, K.D., and Millar, A.J. (2005). Bayesian Clustering and Model Exploration. (Coventry, UK: University of Warwick).
-
- Bastow, R., Mylne, J.S., Lister, C., Lippman, Z., Martienssen, R.A., and Dean, C. (2004). Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 427 164–167. - PubMed
-
- Berriz, G.F., King, O.D., Bryant, B., Sander, C., and Roth, F.P. (2003). Characterizing gene sets with FuncAssociate. Bioinformatics 19 2502–2504. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
