

Mouse-adapted scrapie infection of SN56 cells: greater efficiency with microsome-associated versus purified PrP-res
- PMID: 16474119
- PMCID: PMC1395383
- DOI: 10.1128/JVI.80.5.2106-2117.2006
Mouse-adapted scrapie infection of SN56 cells: greater efficiency with microsome-associated versus purified PrP-res
Abstract
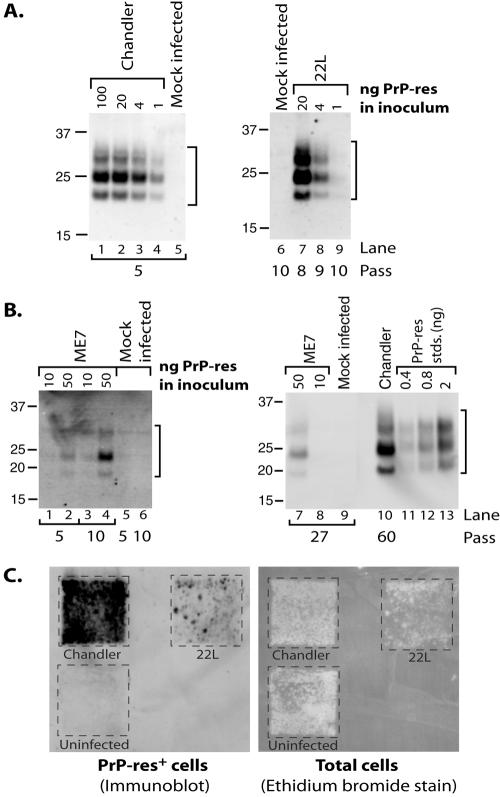
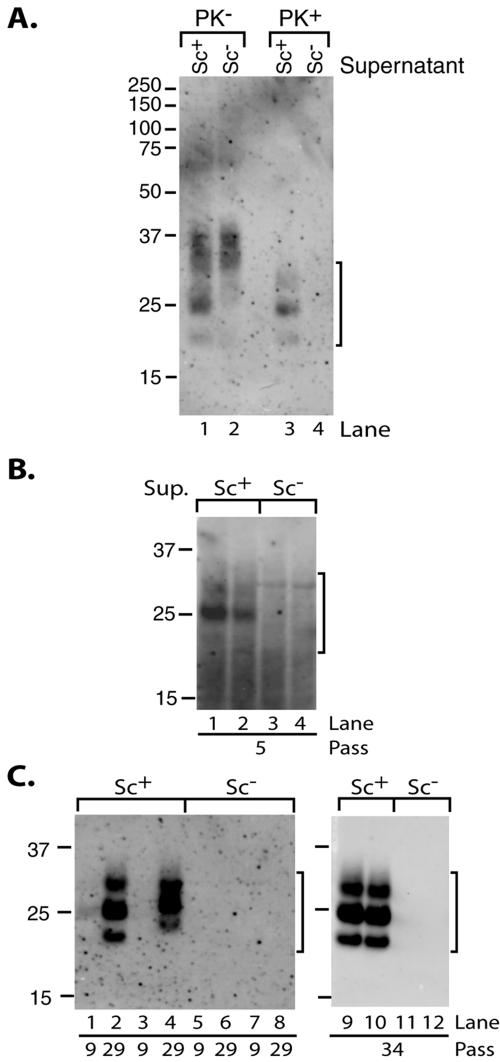
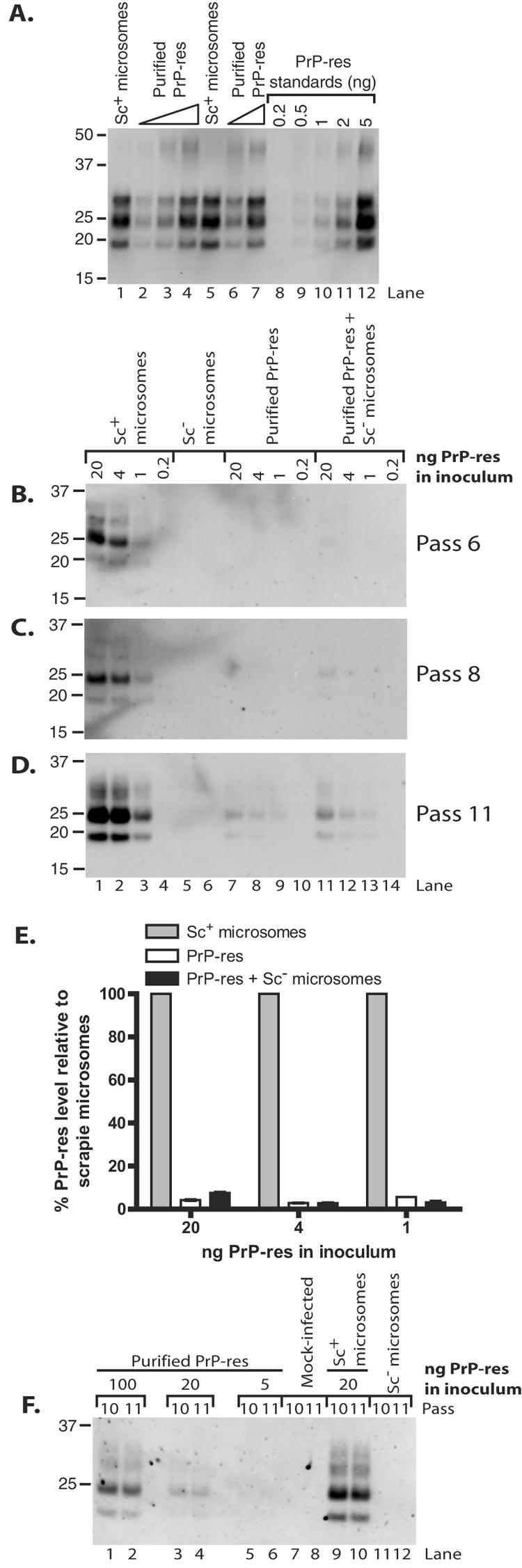
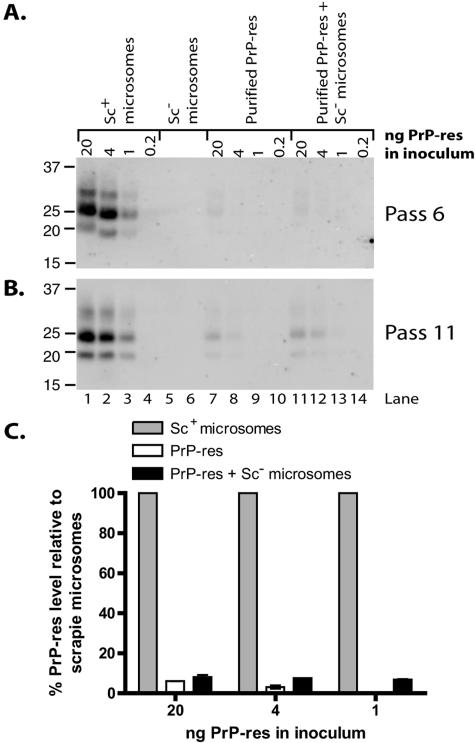
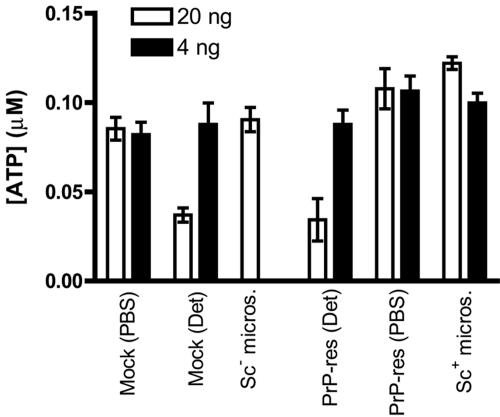
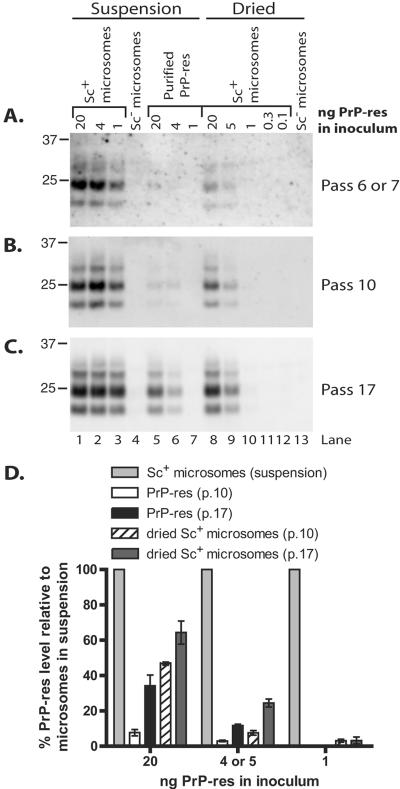
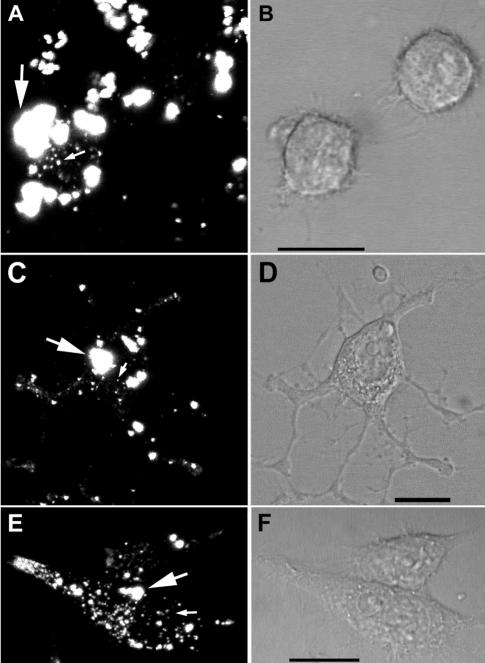
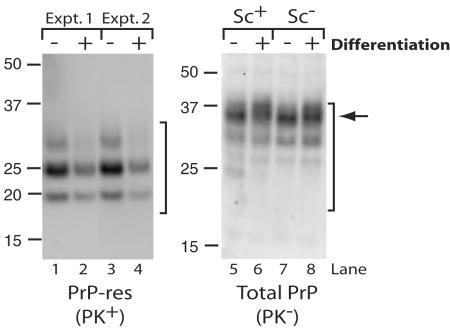
The process by which transmissible spongiform encephalopathy agents, or prions, infect cells is unknown. We employed a new differentiable cell line (SN56) susceptible to infection with three mouse-adapted scrapie strains to gain insight into the cellular infection process. The effect of disease-associated PrP (PrP-res) association with microsomal membranes on infection efficiency was examined by comparing sustained PrP-res production in cells treated with either scrapie brain microsomes or purified, detergent-extracted PrP-res. When normalized for quantity of input PrP-res, scrapie brain microsomes induced dramatically enhanced persistent PrP-res formation compared to purified PrP-res. Infected SN56 cells released low levels of PrP-res into the culture supernatant, which also efficiently initiated infection in recipient cells. Interestingly, microsomes labeled with a fluorescent marker were internalized by SN56 cells in small vesicles, which were subsequently found in neuritic processes. When bound to culture wells to reduce internalization during the infection process, scrapie microsomes induced less long-term PrP-res production than suspended microsomes. Long-term differentiation of infected SN56 cells was accompanied by a decrease in PrP-res formation. Our observations provide evidence that infection of cells is aided by the association of PrP-res with membranes and/or other microsomal constituents.
Figures

References
-
- Aguzzi, A., F. L. Heppner, M. Heikenwalder, M. Prinz, K. Mertz, H. Seeger, and M. Glatzel. 2003. Immune system and peripheral nerves in propagation of prions to CNS. Br. Med. Bull. 66:141-159. - PubMed
-
- Angrand, M., A. Briolay, F. Ronzon, and B. Roux. 1997. Detergent-mediated reconstitution of a glycosyl-phosphatidylinositol-protein into liposomes. Eur. J. Biochem. 250:168-176. - PubMed
-
- Barbosa, J., Jr., A. R. Massensini, M. S. Santos, S. I. Meireles, R. S. Gomez, M. V. Gomez, M. A. Romano-Silva, V. F. Prado, and M. A. Prado. 1999. Expression of the vesicular acetylcholine transporter, proteins involved in exocytosis, and functional calcium signaling in varicosities and soma of a murine septal cell line. J. Neurochem. 73:1881-1893. - PubMed
-
- Baron, G. S., and B. Caughey. 2003. Effect of glycosylphosphatidylinositol anchor-dependent and -independent prion protein association with model raft membranes on conversion to the protease-resistant isoform. J. Biol. Chem. 278:14883-14892. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
