

Strand-biased defect in C/G transversions in hypermutating immunoglobulin genes in Rev1-deficient mice
- PMID: 16476771
- PMCID: PMC2118202
- DOI: 10.1084/jem.20052227
Strand-biased defect in C/G transversions in hypermutating immunoglobulin genes in Rev1-deficient mice
Abstract
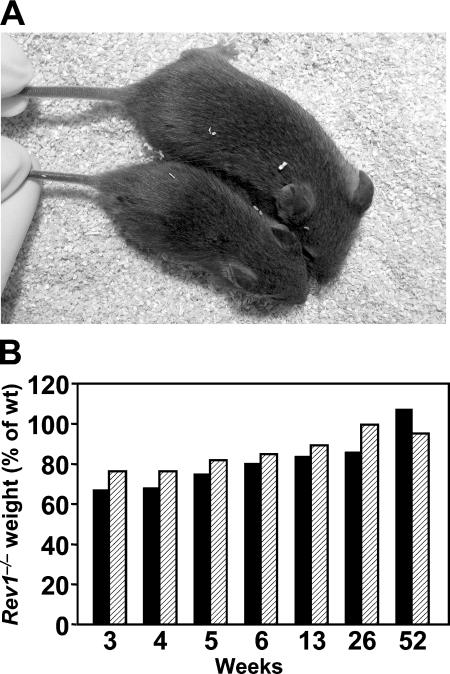
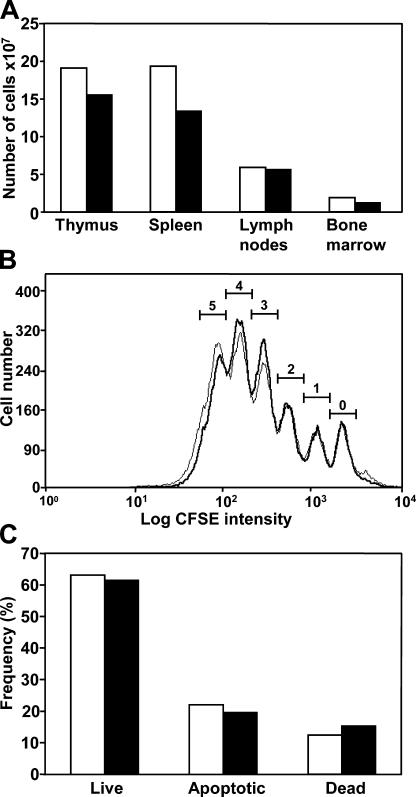
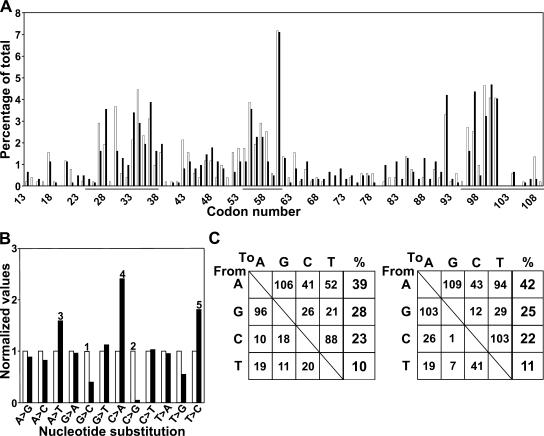
Somatic hypermutation of Ig genes enables B cells of the germinal center to generate high-affinity immunoglobulin variants. Key intermediates in somatic hypermutation are deoxyuridine lesions, introduced by activation-induced cytidine deaminase. These lesions can be processed further to abasic sites by uracil DNA glycosylase. Mutagenic replication of deoxyuridine, or of its abasic derivative, by translesion synthesis polymerases is hypothesized to underlie somatic hypermutation. Rev1 is a translesion synthesis polymerase that in vitro incorporates uniquely deoxycytidine opposite deoxyuridine and abasic residues. To investigate a role of Rev1 in mammalian somatic hypermutation we have generated mice deficient for Rev1. Although Rev1-/- mice display transient growth retardation, proliferation of Rev1-/- LPS-stimulated B cells is indistinguishable from wild-type cells. In mutated Ig genes from Rev1-/- mice, C to G transversions were virtually absent in the nontranscribed (coding) strand and reduced in the transcribed strand. This defect is associated with an increase of A to T, C to A, and T to C substitutions. These results indicate that Rev1 incorporates deoxycytidine residues, most likely opposite abasic nucleotides, during somatic hypermutation. In addition, loss of Rev1 causes compensatory increase in mutagenesis by other translesion synthesis polymerases.
Figures
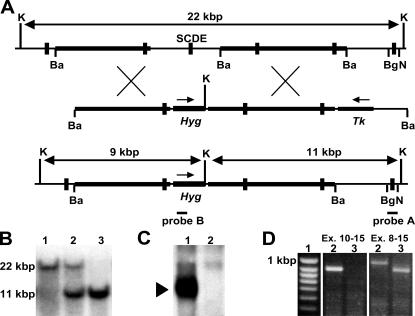
References
-
- Prakash, S., R.E. Johnson, and L. Prakash. 2005. Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function. Annu. Rev. Biochem. 74:317–353. - PubMed
-
- Masuda, Y., M. Takahashi, S. Fukuda, M. Sumii, and K. Kamiya. 2002. Mechanisms of dCMP transferase reactions catalyzed by mouse Rev1 protein. J. Biol. Chem. 277:3040–3046. - PubMed
-
- Gibbs, P.E.M., J. McDonald, R. Woodgate, and C.W. Lawrence. 2005. The relative roles in vivo of Saccharomyces cerevisiae Pol eta, Pol zeta, Rev1 protein and Pol32 in the bypass and mutation induction of an abasic site, T-T (6-4) photoadduct and T-T cis-syn cyclobutane dimer. Genetics. 169:575–582. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
