

Permissive transcriptional activity at the centromere through pockets of DNA hypomethylation
- PMID: 16477312
- PMCID: PMC1361766
- DOI: 10.1371/journal.pgen.0020017
Permissive transcriptional activity at the centromere through pockets of DNA hypomethylation
Abstract
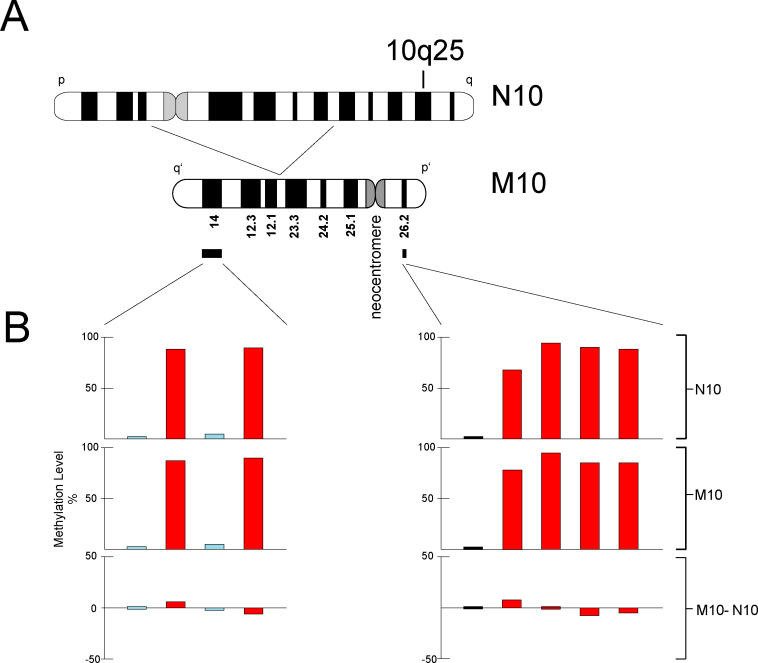
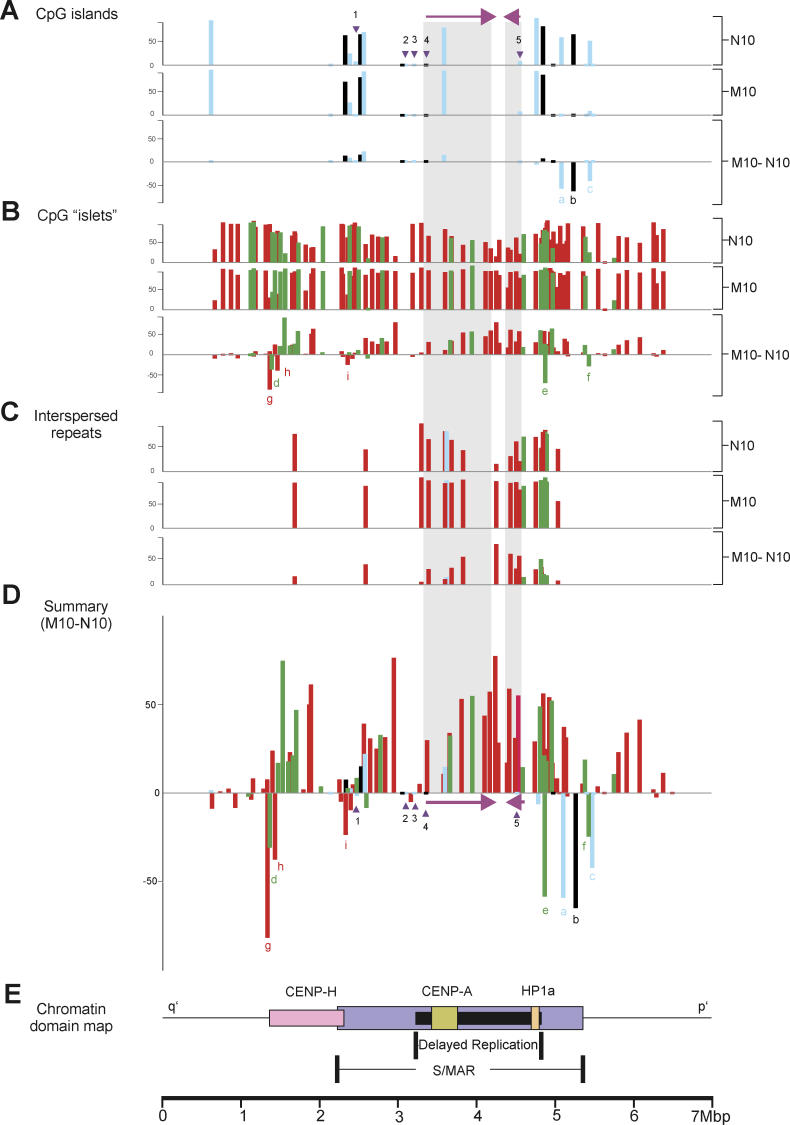
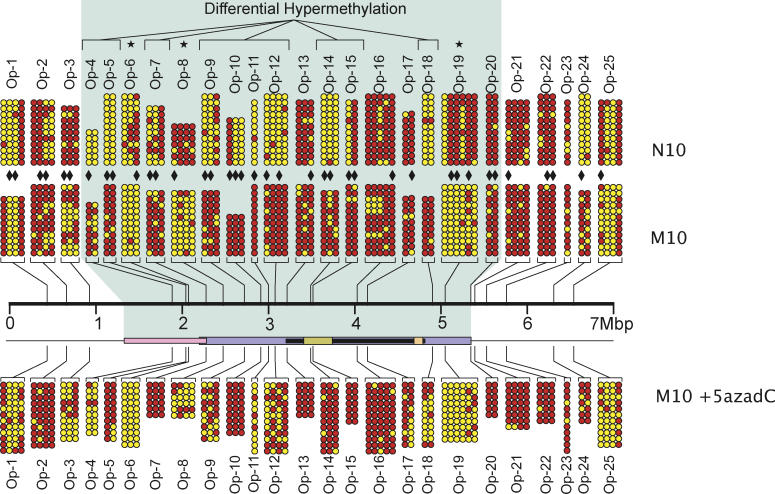
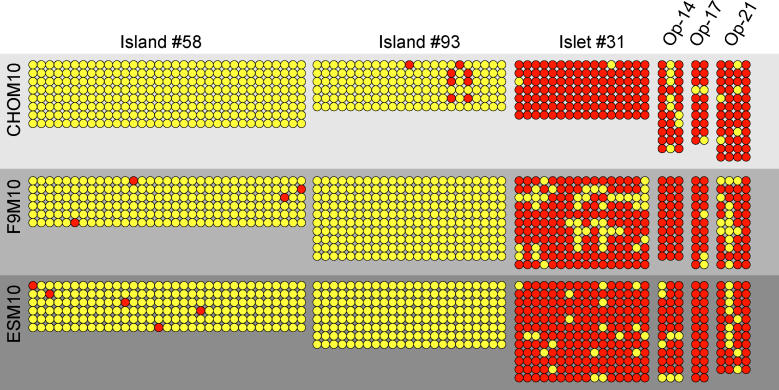
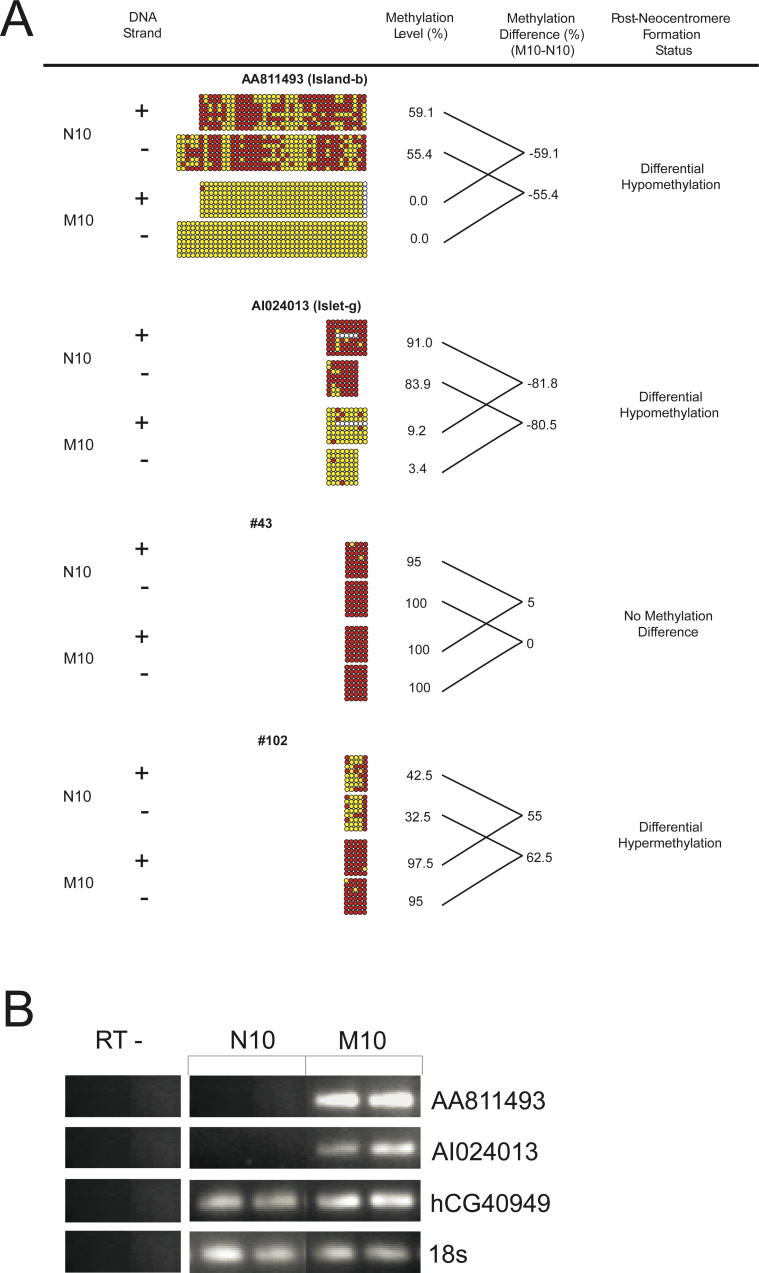
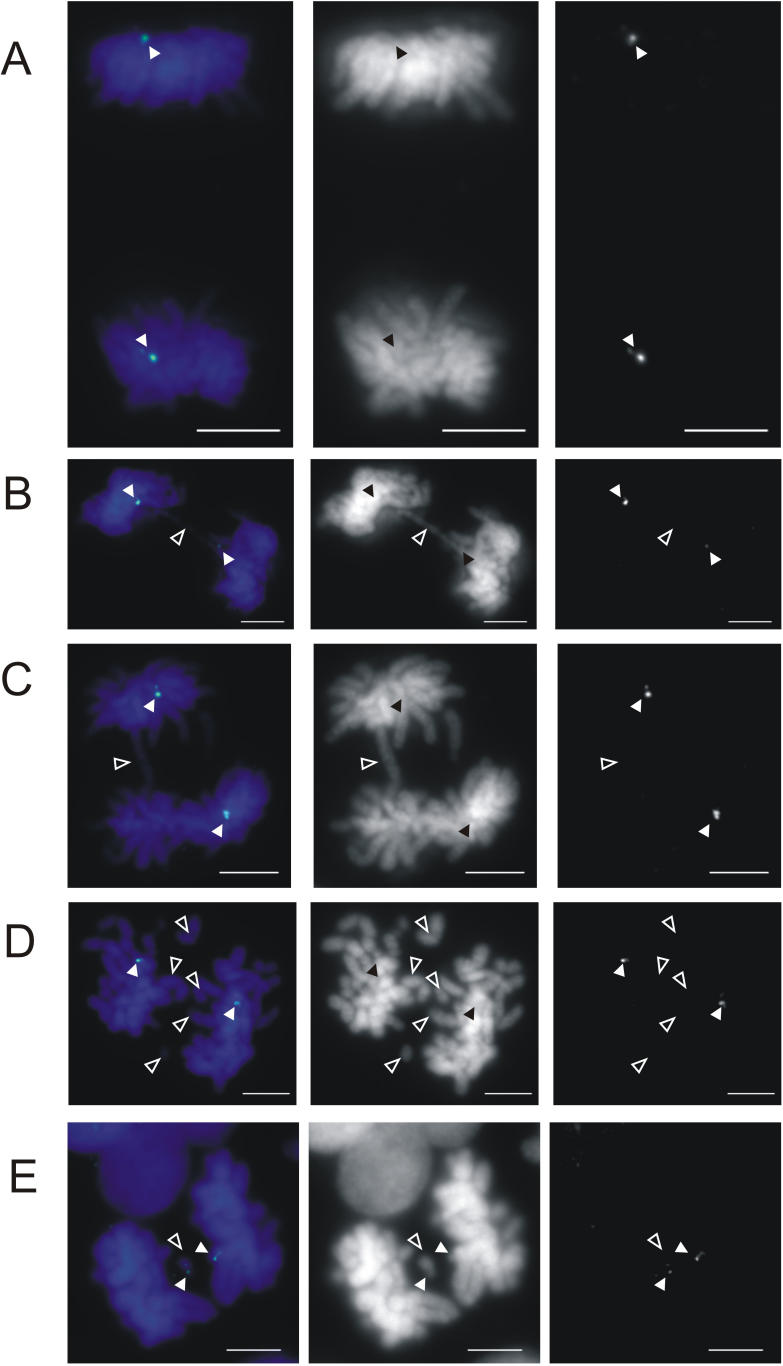
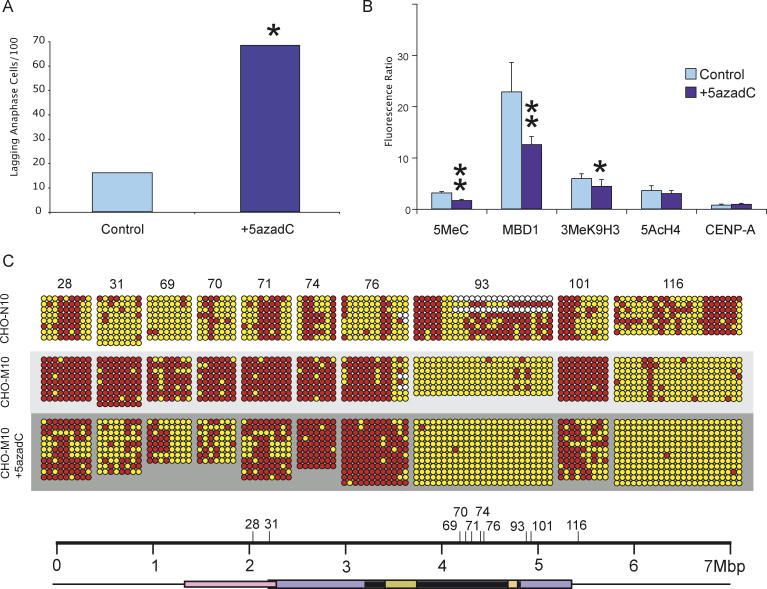
DNA methylation is a hallmark of transcriptional silencing, yet transcription has been reported at the centromere. To address this apparent paradox, we employed a fully sequence-defined ectopic human centromere (or neocentromere) to investigate the relationship between DNA methylation and transcription. We used sodium bisulfite PCR and sequencing to determine the methylation status of 2,041 CpG dinucleotides distributed across a 6.76-Mbp chromosomal region containing a neocentromere. These CpG dinucleotides were associated with conventional and nonconventional CpG islands. We found an overall hypermethylation of the neocentric DNA at nonconventional CpG islands that we designated as CpG islets and CpG orphans. The observed hypermethylation was consistent with the presence of a presumed transcriptionally silent chromatin state at the neocentromere. Within this neocentric chromatin, specific sites of active transcription and the centromeric chromatin boundary are defined by DNA hypomethylation. Our data demonstrate, for the first time to our knowledge, a correlation between DNA methylation and centromere formation in mammals, and that transcription and "chromatin-boundary activity" are permissible at the centromere through the selective hypomethylation of pockets of sequences without compromising the overall silent chromatin state and function of the centromere.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
Similar articles
-
Transcriptional gene silencing promotes DNA hypermethylation through a sequential change in chromatin modifications in cancer cells.Cancer Res. 2004 Jun 1;64(11):3871-7. doi: 10.1158/0008-5472.CAN-03-3690. Cancer Res. 2004. PMID: 15172996
-
Altered chromatin structure associated with methylation-induced gene silencing in cancer cells: correlation of accessibility, methylation, MeCP2 binding and acetylation.Nucleic Acids Res. 2001 Nov 15;29(22):4598-606. doi: 10.1093/nar/29.22.4598. Nucleic Acids Res. 2001. PMID: 11713309 Free PMC article.
-
Satellite 2 methylation patterns in normal and ICF syndrome cells and association of hypomethylation with advanced replication.Hum Genet. 2001 Oct;109(4):452-62. doi: 10.1007/s004390100590. Hum Genet. 2001. PMID: 11702227
-
CpG islands in chromatin organization and gene expression.J Biochem. 1999 Feb;125(2):217-22. doi: 10.1093/oxfordjournals.jbchem.a022276. J Biochem. 1999. PMID: 9990116 Review.
-
DNA methylation and chromatin structure: the puzzling CpG islands.J Cell Biochem. 2005 Feb 1;94(2):257-65. doi: 10.1002/jcb.20325. J Cell Biochem. 2005. PMID: 15546139 Review.
Cited by
-
Efficient neocentromere formation is suppressed by gene conversion to maintain centromere function at native physical chromosomal loci in Candida albicans.Genome Res. 2013 Apr;23(4):638-52. doi: 10.1101/gr.141614.112. Epub 2013 Feb 25. Genome Res. 2013. PMID: 23439889 Free PMC article.
-
Epigenetic Regulation of Centromere Chromatin Stability by Dietary and Environmental Factors.Adv Nutr. 2017 Nov 15;8(6):889-904. doi: 10.3945/an.117.016402. Print 2017 Nov. Adv Nutr. 2017. PMID: 29141972 Free PMC article. Review.
-
Formation of a functional maize centromere after loss of centromeric sequences and gain of ectopic sequences.Plant Cell. 2013 Jun;25(6):1979-89. doi: 10.1105/tpc.113.110015. Epub 2013 Jun 14. Plant Cell. 2013. PMID: 23771890 Free PMC article.
-
Asymmetric distribution of gene expression in the centromeric region of rice chromosome 5.Front Plant Sci. 2011 Jun 6;2:16. doi: 10.3389/fpls.2011.00016. eCollection 2011. Front Plant Sci. 2011. PMID: 22639581 Free PMC article.
-
Keeping the Centromere under Control: A Promising Role for DNA Methylation.Cells. 2019 Aug 16;8(8):912. doi: 10.3390/cells8080912. Cells. 2019. PMID: 31426433 Free PMC article. Review.
References
-
- Choo KHA. Domain organization at the centromere and neocentromere. Dev Cell. 2001;1:165–177. - PubMed
-
- Pidoux AL, Allshire RC. Kinetochore and heterochromatin domains of the fission yeast centromere. Chromosome Res. 2004;12:521–534. - PubMed
-
- Amor DJ, Kalitsis P, Sumer H, Choo KHA. Building the centromere: From foundation proteins to 3D organization. Trends Cell Biol. 2004;14:359–368. - PubMed
-
- Cleveland DW, Mao Y, Sullivan KF. Centromeres and kinetochores: From epigenetics to mitotic checkpoint signaling. Cell. 2003;112:407–421. - PubMed
-
- Craig JM, Earle E, Canham P, Wong LH, Anderson M, et al. Analysis of mammalian proteins involved in chromatin modification reveals new metaphase centromeric proteins and distinct chromosomal distribution patterns. Hum Mol Genet. 2003;12:3109–3121. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
