

Regulation of human neural precursor cells by laminin and integrins
- PMID: 16477652
- PMCID: PMC2409144
- DOI: 10.1002/jnr.20778
Regulation of human neural precursor cells by laminin and integrins
Abstract
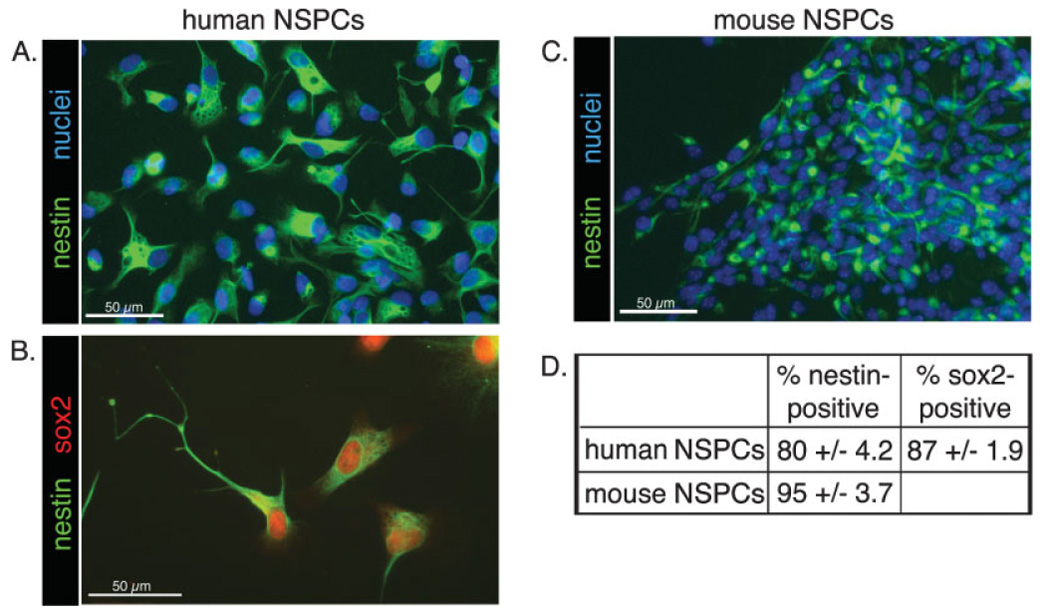
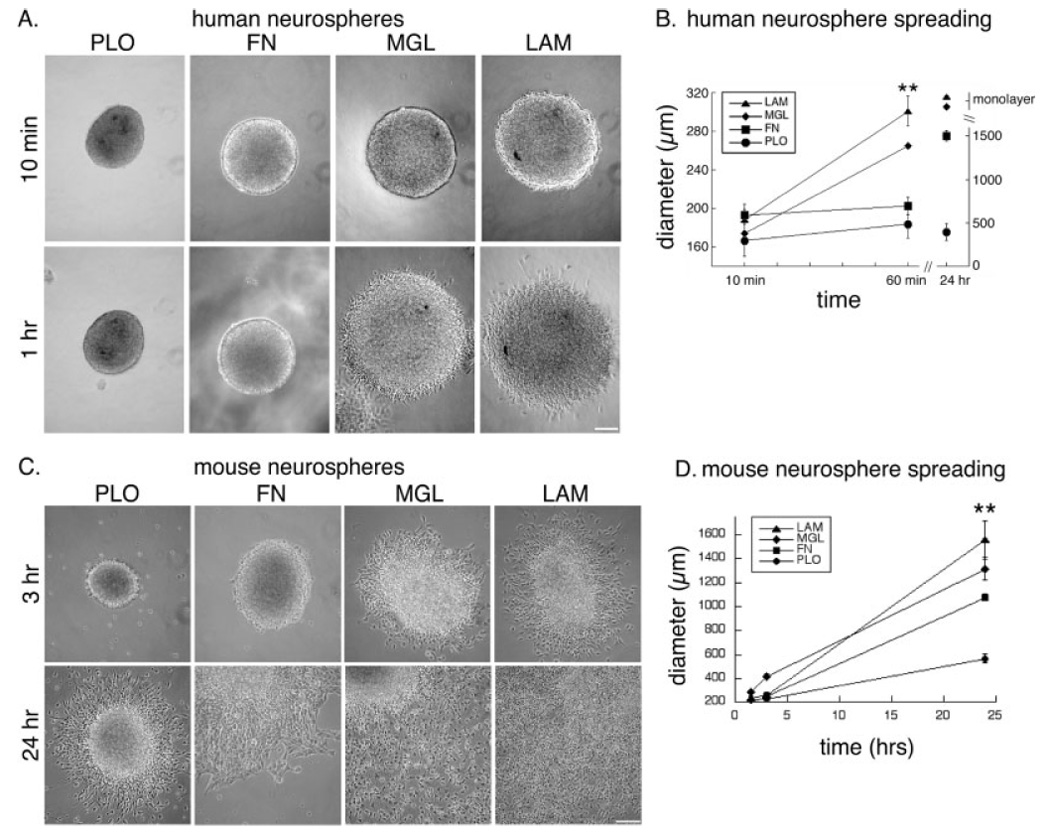
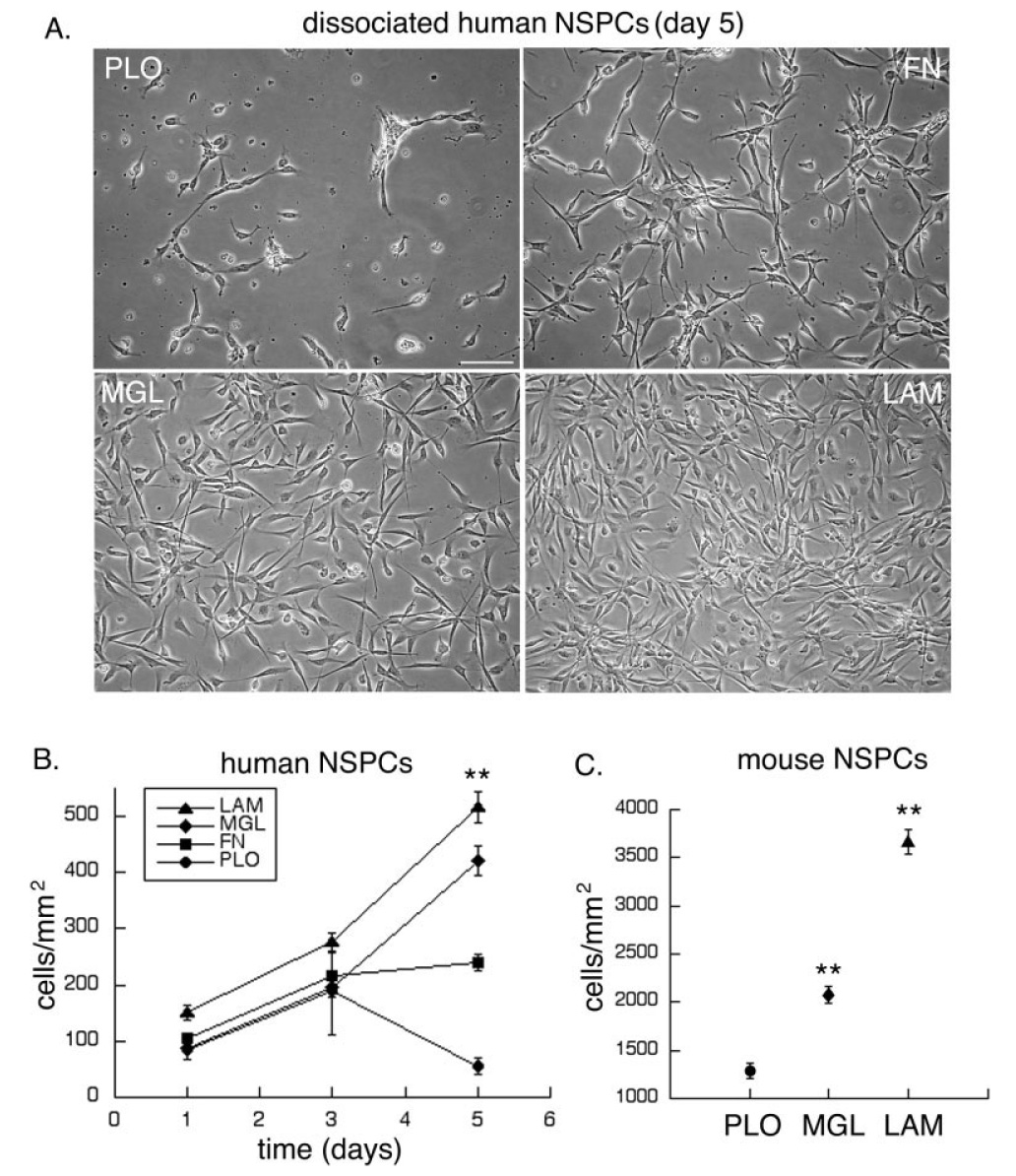
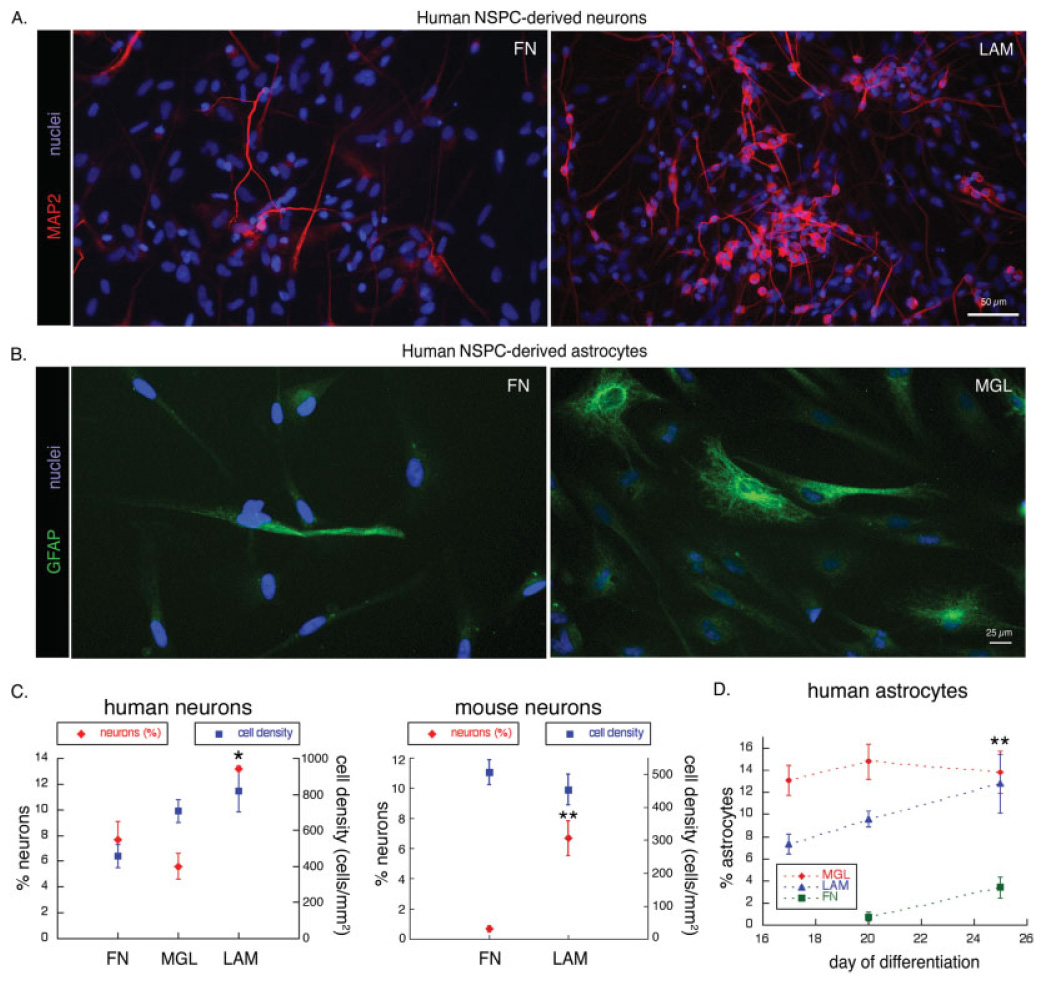
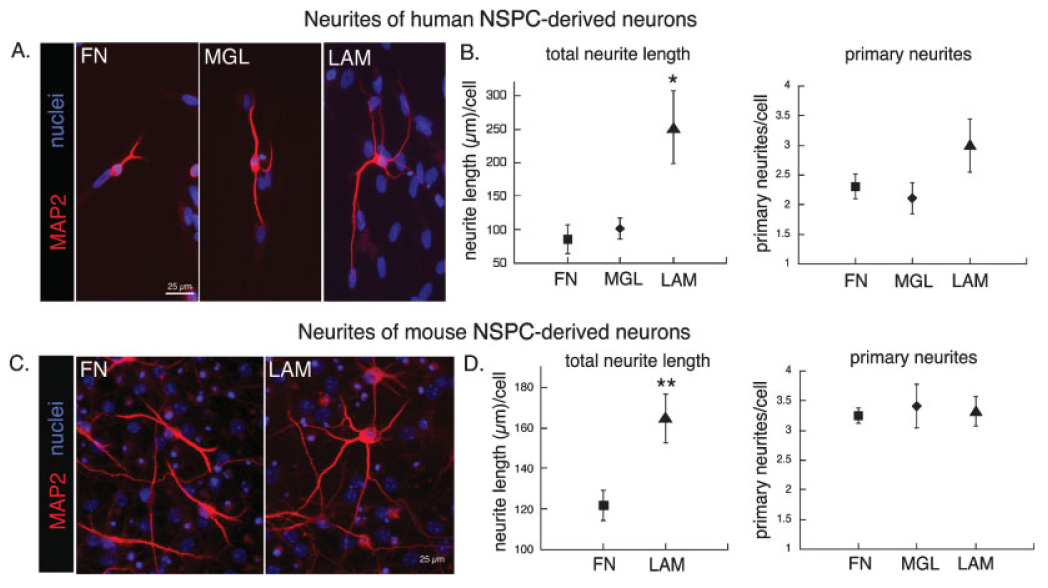
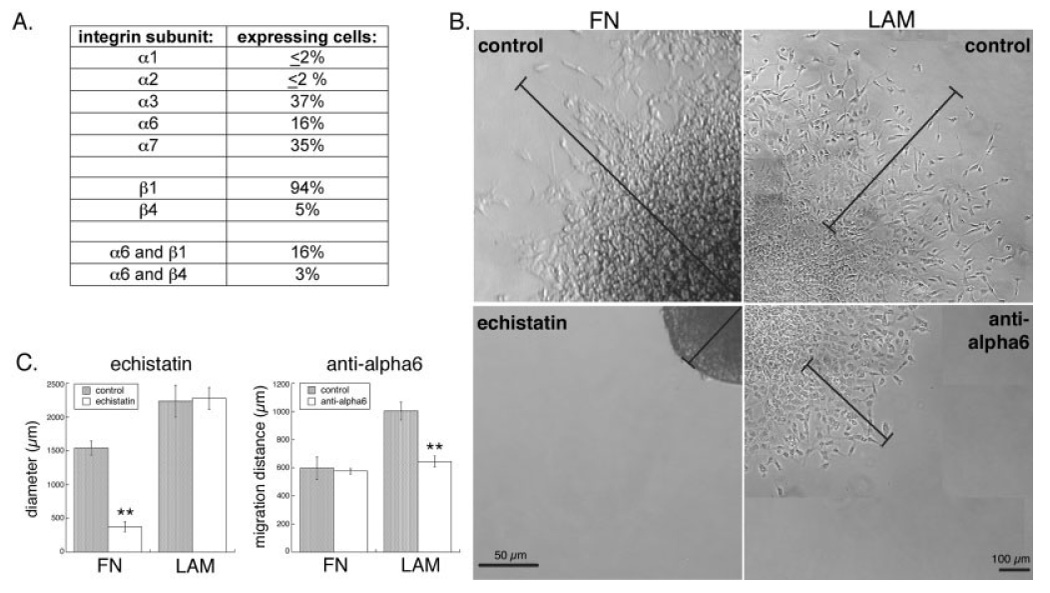
Deciphering the factors that regulate human neural stem cells will greatly aid in their use as models of development and as therapeutic agents. The extracellular matrix (ECM) is a component of stem cell niches in vivo and regulates multiple functions in diverse cell types, yet little is known about its effects on human neural stem/precursor cells (NSPCs). We therefore plated human NSPCs on four different substrates (poly-L-ornithine, fibronectin, laminin, and matrigel) and compared their responses with those of mouse NSPCs. Compared with the other substrates, laminin matrices enhanced NSPC migration, expansion, differentiation into neurons and astrocytes, and elongation of neurites from NSPC-derived neurons. Laminin had a similar spectrum of effects on both human and mouse cells, highlighting the evolutionary conservation of NSPC regulation by this component of the ECM. Flow cytometry revealed that human NSPCs express on their cell surfaces the laminin-binding integrins alpha3, alpha6, alpha7, beta1, and beta4, and function-blocking antibodies to the alpha6 subunit confirmed a role for integrins in laminin-dependent migration of human NSPCs. These results define laminin and its integrin receptors as key regulators of human NSPCs.
Figures
References
-
- Alvarez-Buylla A, Lim DA. For the long run: maintaining germinal niches in the adult brain. Neuron. 2004;41:683–686. - PubMed
-
- Anlar B, Atilla P, Cakar AN, Kose MF, Beksac MS, Dagdeviren A, Akcoren Z. Expression of adhesion and extracellular matrix molecules in the developing human brain. J Child Neurol. 2002;17:707–713. - PubMed
-
- Bokel C, Brown NH. Integrins in development: moving on, responding to, and sticking to the extracellular matrix. Dev Cell. 2002;3:311–321. - PubMed
-
- Burgeson RE, Chiquet M, Deutzmann R, Ekblom P, Engel J, Kleinman H, Martin GR, Meneguzzi G, Paulsson M, Sanes J, Timpl R, Tryggvason K, Yamada Y, Yurchenco PD. A new nomenclature for the laminins. Matrix Biol. 1994;14:209–211. - PubMed
-
- Cai J, Wu Y, Mirua T, Pierce JL, Lucero MT, Albertine KH, Spangrude GJ, Rao MS. Properties of a fetal multipotent neural stem cell (NEP cell) Dev Biol. 2002;251:221–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
