

Gadd34 requirement for normal hemoglobin synthesis
- PMID: 16478986
- PMCID: PMC1430266
- DOI: 10.1128/MCB.26.5.1644-1653.2006
Gadd34 requirement for normal hemoglobin synthesis
Abstract
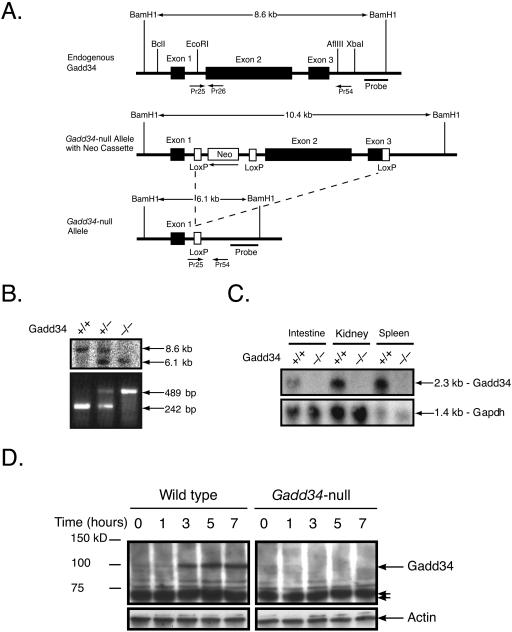
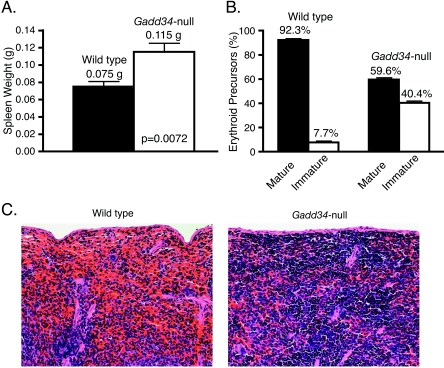
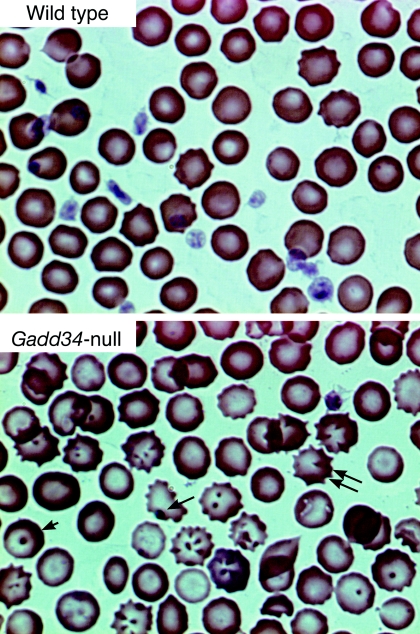
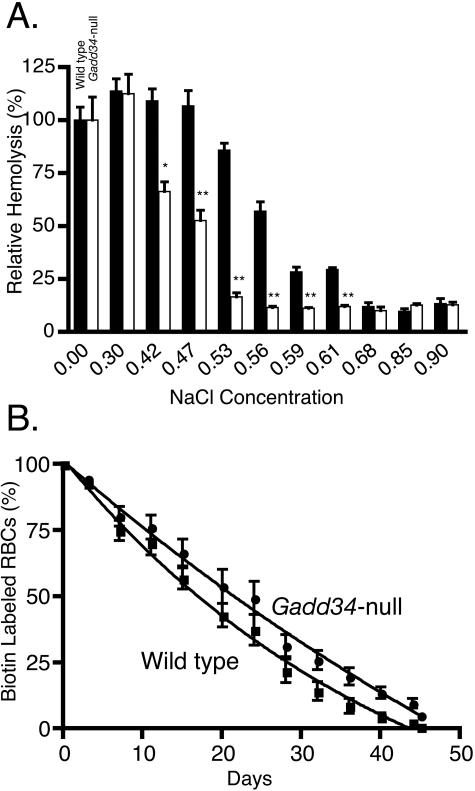
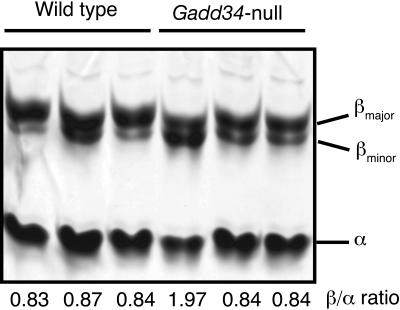
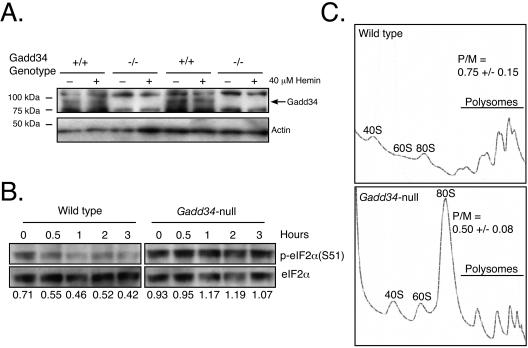
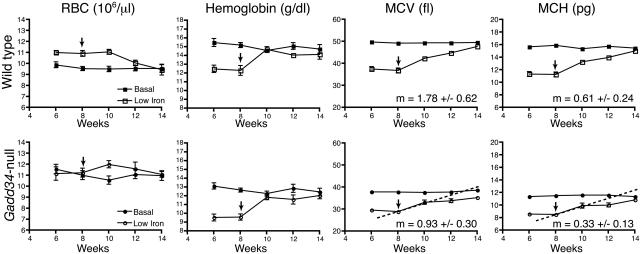
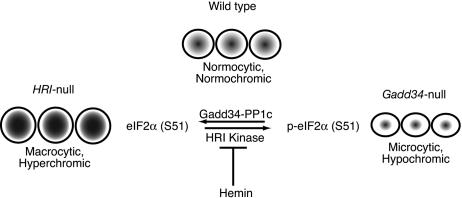
The protein encoded by growth arrest and DNA damage-inducible transcript 34 (Gadd34) is associated with translation initiation regulation following certain stress responses. Through interaction with the protein phosphatase 1 catalytic subunit (PP1c), Gadd34 recruits PP1c for the removal of an inhibitory phosphate group on the alpha subunit of elongation initiation factor 2, thereby reversing the shutoff of protein synthesis initiated by stress-inducible kinases. In the absence of stress, the physiologic consequences of Gadd34 function are not known. Initial analysis of Gadd34-null mice revealed several significant findings, including hypersplenism, decreased erythrocyte volume, increased numbers of circulating erythrocytes, and decreased hemoglobin content, resembling some thalassemia syndromes. Biochemical analysis of the hemoglobin-producing reticulocyte (an erythrocyte precursor) revealed that the decreased hemoglobin content in the Gadd34-null erythrocyte is due to the reduced initiation of the globin translation machinery. We propose that an equilibrium state exists between Gadd34/PP1c and the opposing heme-regulated inhibitor kinase during hemoglobin synthesis in the reticulocyte.
Figures
Similar articles
-
Growth arrest and DNA damage-inducible protein GADD34 targets protein phosphatase 1 alpha to the endoplasmic reticulum and promotes dephosphorylation of the alpha subunit of eukaryotic translation initiation factor 2.Mol Cell Biol. 2003 Feb;23(4):1292-303. doi: 10.1128/MCB.23.4.1292-1303.2003. Mol Cell Biol. 2003. PMID: 12556489 Free PMC article.
-
GADD34-PP1c recruited by Smad7 dephosphorylates TGFbeta type I receptor.J Cell Biol. 2004 Jan 19;164(2):291-300. doi: 10.1083/jcb.200307151. Epub 2004 Jan 12. J Cell Biol. 2004. PMID: 14718519 Free PMC article.
-
Growth arrest and DNA damage-inducible protein GADD34 assembles a novel signaling complex containing protein phosphatase 1 and inhibitor 1.Mol Cell Biol. 2001 Oct;21(20):6841-50. doi: 10.1128/MCB.21.20.6841-6850.2001. Mol Cell Biol. 2001. PMID: 11564868 Free PMC article.
-
Disruption of the PP1/GADD34 complex induces calreticulin exposure.Cell Cycle. 2009 Dec;8(23):3971-7. doi: 10.4161/cc.8.23.10191. Epub 2009 Dec 28. Cell Cycle. 2009. PMID: 19901557
-
Stress-induced gene expression requires programmed recovery from translational repression.EMBO J. 2003 Mar 3;22(5):1180-7. doi: 10.1093/emboj/cdg112. EMBO J. 2003. PMID: 12606582 Free PMC article.
Cited by
-
Raphin-1 mediates the survival and sensitivity to radiation of pediatric-type diffuse high-grade glioma via phosphorylated eukaryotic initiation factor 2α-dependent and -independent processes.Mol Oncol. 2025 Sep;19(9):2648-2669. doi: 10.1002/1878-0261.70081. Epub 2025 Jul 9. Mol Oncol. 2025. PMID: 40632659 Free PMC article.
-
The mTORC1/4E-BP pathway coordinates hemoglobin production with L-leucine availability.Sci Signal. 2015 Apr 14;8(372):ra34. doi: 10.1126/scisignal.aaa5903. Sci Signal. 2015. PMID: 25872869 Free PMC article.
-
RNA Binding Proteins and Regulation of mRNA Translation in Erythropoiesis.Front Physiol. 2018 Jul 24;9:910. doi: 10.3389/fphys.2018.00910. eCollection 2018. Front Physiol. 2018. PMID: 30087616 Free PMC article. Review.
-
Complementary Roles of GADD34- and CReP-Containing Eukaryotic Initiation Factor 2α Phosphatases during the Unfolded Protein Response.Mol Cell Biol. 2016 Jun 15;36(13):1868-80. doi: 10.1128/MCB.00190-16. Print 2016 Jul 1. Mol Cell Biol. 2016. PMID: 27161320 Free PMC article.
-
Functions and therapeutic potential of protein phosphatase 1: Insights from mouse genetics.Biochim Biophys Acta Mol Cell Res. 2019 Jan;1866(1):16-30. doi: 10.1016/j.bbamcr.2018.07.019. Epub 2018 Jul 26. Biochim Biophys Acta Mol Cell Res. 2019. PMID: 30056088 Free PMC article. Review.
References
-
- Alter, B. P., S. C. Goff, G. D. Efremov, M. E. Gravely, and T. H. Huisman. 1980. Globin chain electrophoresis: a new approach to the determination of the G gamma/A gamma ratio in fetal haemoglobin and to studies of globin synthesis. Br. J. Haematol. 44:527-534. - PubMed
-
- Bauer, B. N., M. Rafie-Kolpin, L. Lu, A. Han, and J. J. Chen. 2001. Multiple autophosphorylation is essential for the formation of the active and stable homodimer of heme-regulated eIF2alpha kinase. Biochemistry 40:11543-11551. - PubMed
-
- Brush, M. H., D. C. Weiser, and S. Shenolikar. 2003. Growth arrest and DNA damage-inducible protein GADD34 targets protein phosphatase 1α to the endoplasmic reticulum and promotes dephosphorylation of the α subunit of eukaryotic translation initiation factor 2. Mol. Cell. Biol. 23:1292-1303. - PMC - PubMed
-
- Chefalo, P. J., J. Oh, M. Rafie-Kolpin, B. Kan, and J. J. Chen. 1998. Heme-regulated eIF-2alpha kinase purifies as a hemoprotein. Eur. J. Biochem. 258:820-830. - PubMed
-
- Chomczynski, P., and N. Sacchi. 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 162:156-159. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases