

Matrix metalloproteinase-9 is required for hippocampal late-phase long-term potentiation and memory
- PMID: 16481424
- PMCID: PMC4428329
- DOI: 10.1523/JNEUROSCI.4359-05.2006
Matrix metalloproteinase-9 is required for hippocampal late-phase long-term potentiation and memory
Abstract
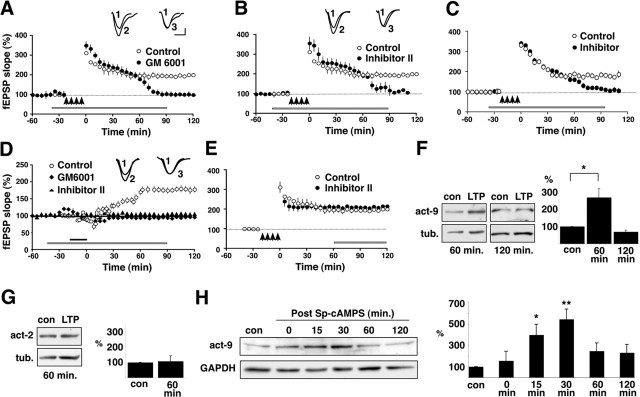
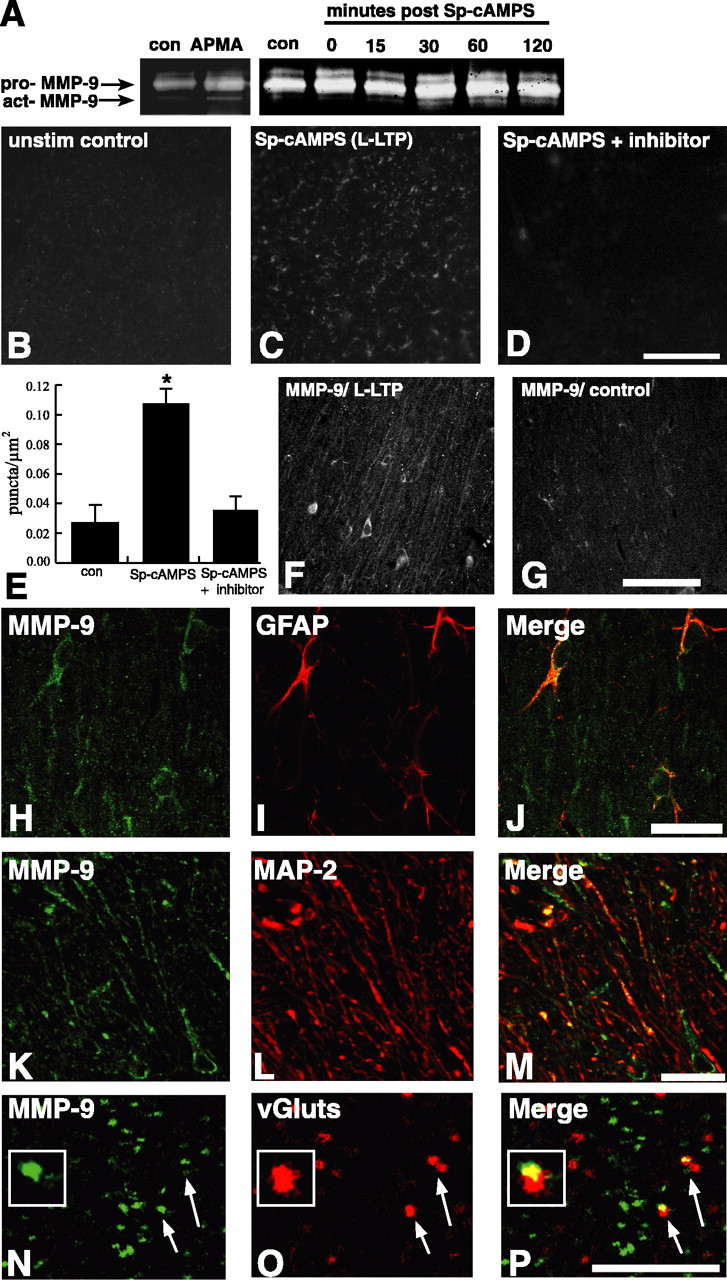
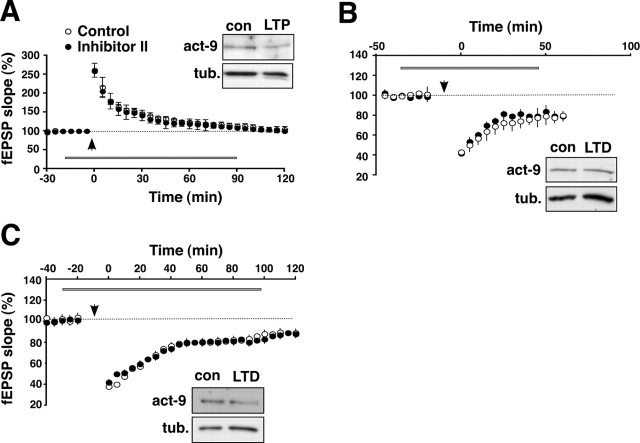
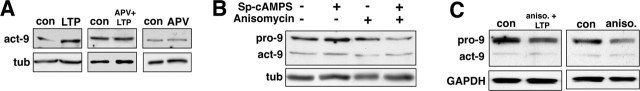
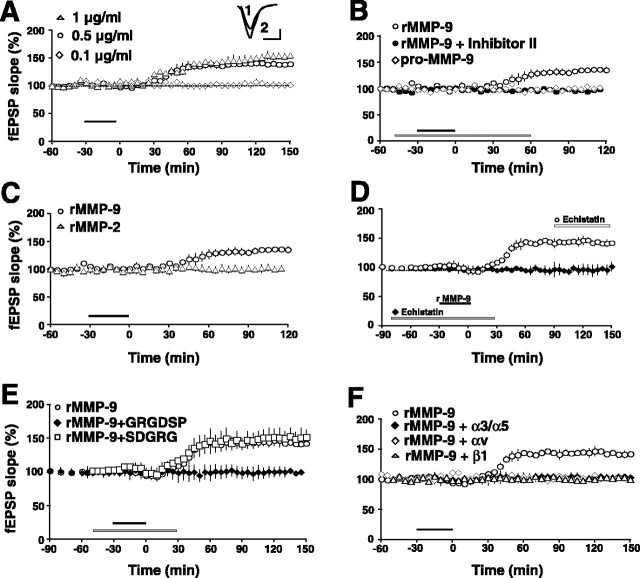
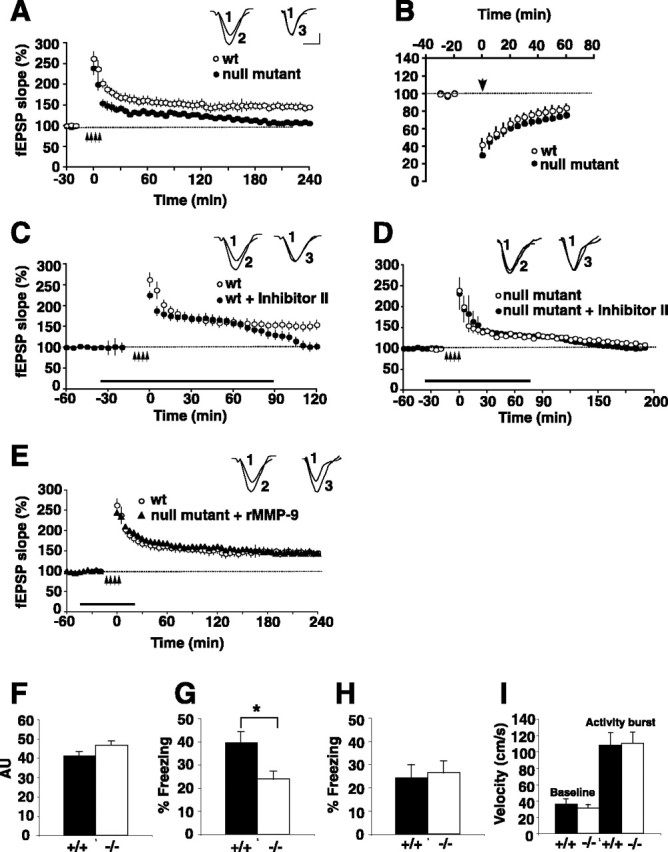
Matrix metalloproteinases (MMPs) are extracellular proteases that have well recognized roles in cell signaling and remodeling in many tissues. In the brain, their activation and function are customarily associated with injury or pathology. Here, we demonstrate a novel role for MMP-9 in hippocampal synaptic physiology, plasticity, and memory. MMP-9 protein levels and proteolytic activity are rapidly increased by stimuli that induce late-phase long-term potentiation (L-LTP) in area CA1. Such regulation requires NMDA receptors and protein synthesis. Blockade of MMP-9 pharmacologically prevents induction of L-LTP selectively; MMP-9 plays no role in, nor is regulated during, other forms of short-term synaptic potentiation or long-lasting synaptic depression. Similarly, in slices from MMP-9 null-mutant mice, hippocampal LTP, but not long-term depression, is impaired in magnitude and duration; adding recombinant active MMP-9 to null-mutant slices restores the magnitude and duration of LTP to wild-type levels. Activated MMP-9 localizes in part to synapses and modulates hippocampal synaptic physiology through integrin receptors, because integrin function-blocking reagents prevent an MMP-9-mediated potentiation of synaptic signal strength. The fundamental importance of MMP-9 function in modulating hippocampal synaptic physiology and plasticity is underscored by behavioral impairments in hippocampal-dependent memory displayed by MMP-9 null-mutant mice. Together, these data reveal new functions for MMPs in synaptic and behavioral plasticity.
Figures
References
-
- Arai K, Lee SR, Lo EH (2003). Essential role for ERK mitogen-activated protein kinase in matrix metalloproteinase-9 regulation in rat cortical astrocytes. Glia 43:254–264. - PubMed
-
- Baranes D, Lederfein D, Huang YY, Chen M, Bailey CH, Kandel ER (1998). Tissue plasminogen activator contributes to the late phase of LTP and to synaptic growth in the hippocampal mossy fiber pathway. Neuron 21:813–825. - PubMed
-
- Barria A, Malinow R (2005). NMDA receptor subunit composition controls synaptic plasticity by regulating binding to CaMKII. Neuron 48:289–301. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous