

Polycomb group-dependent Cyclin A repression in Drosophila
- PMID: 16481477
- PMCID: PMC1369051
- DOI: 10.1101/gad.357106
Polycomb group-dependent Cyclin A repression in Drosophila
Abstract
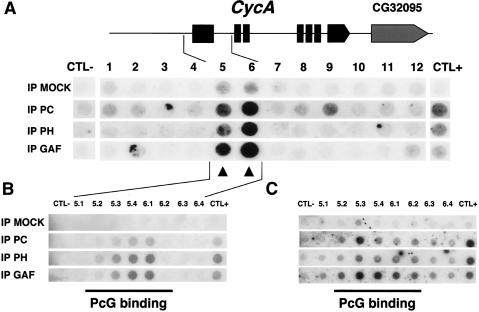
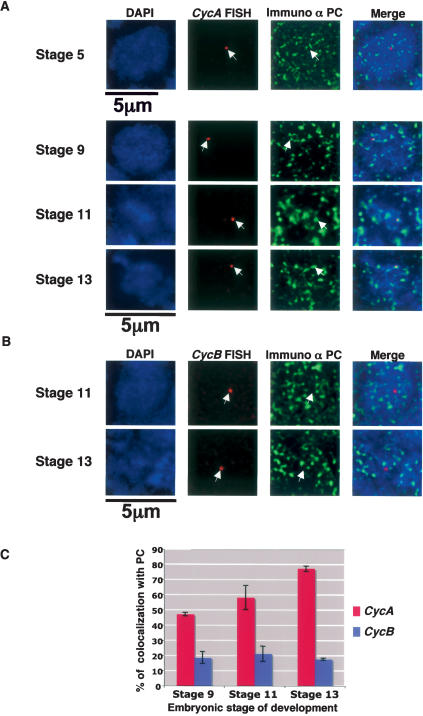
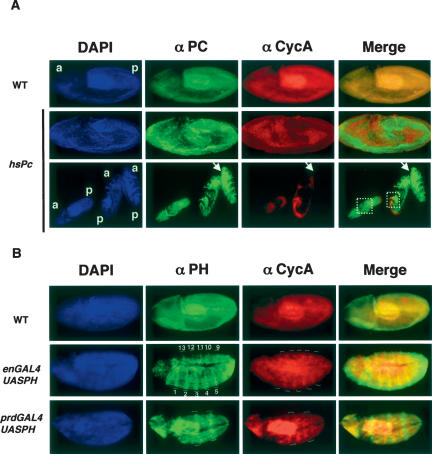
Polycomb group (PcG) and trithorax group (trxG) proteins are well known for their role in the maintenance of silent and active expression states of homeotic genes. However, PcG proteins may also be required for the control of cellular proliferation in vertebrates. In Drosophila, PcG factors act by associating with specific DNA regions termed PcG response elements (PREs). Here, we have investigated whether Drosophila cell cycle genes are directly regulated by PcG proteins through PREs. We have isolated a PRE that regulates Cyclin A (CycA) expression. This sequence is bound by the Polycomb (PC) and Polyhomeotic (PH) proteins of the PcG, and also by GAGA factor (GAF), a trxG protein that is usually found associated with PREs. This sequence causes PcG- and trxG-dependent variegation of the mini-white reporter gene in transgenic flies. The combination of FISH with PC immunostaining in embryonic cells shows that the endogenous CycA gene colocalizes with PC at foci of high PC concentration named PcG bodies. Finally, loss of function of the Pc gene and overexpression of Pc and ph trigger up-regulation and down-regulation, respectively, of CycA expression in embryos. These results demonstrate that CycA is directly regulated by PcG proteins, linking them to cell cycle control in vivo.
Figures




References
-
- Beuchle D., Struhl, G., and Müller, J. 2001. Polycomb group proteins and heritable silencing of Drosophila Hox genes. Development 128: 993–1004. - PubMed
-
- Brand A.H. and Perrimon, N. 1993. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118: 401–415. - PubMed
-
- Brehm A. and Kouzarides, T. 1999. Retinoblastoma protein meets chromatin. Trends Biochem. Sci. 24: 142–145. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases