

Impact of behavior on central and peripheral circadian clocks in the common vole Microtus arvalis, a mammal with ultradian rhythms
- PMID: 16481616
- PMCID: PMC1413878
- DOI: 10.1073/pnas.0507825103
Impact of behavior on central and peripheral circadian clocks in the common vole Microtus arvalis, a mammal with ultradian rhythms
Abstract
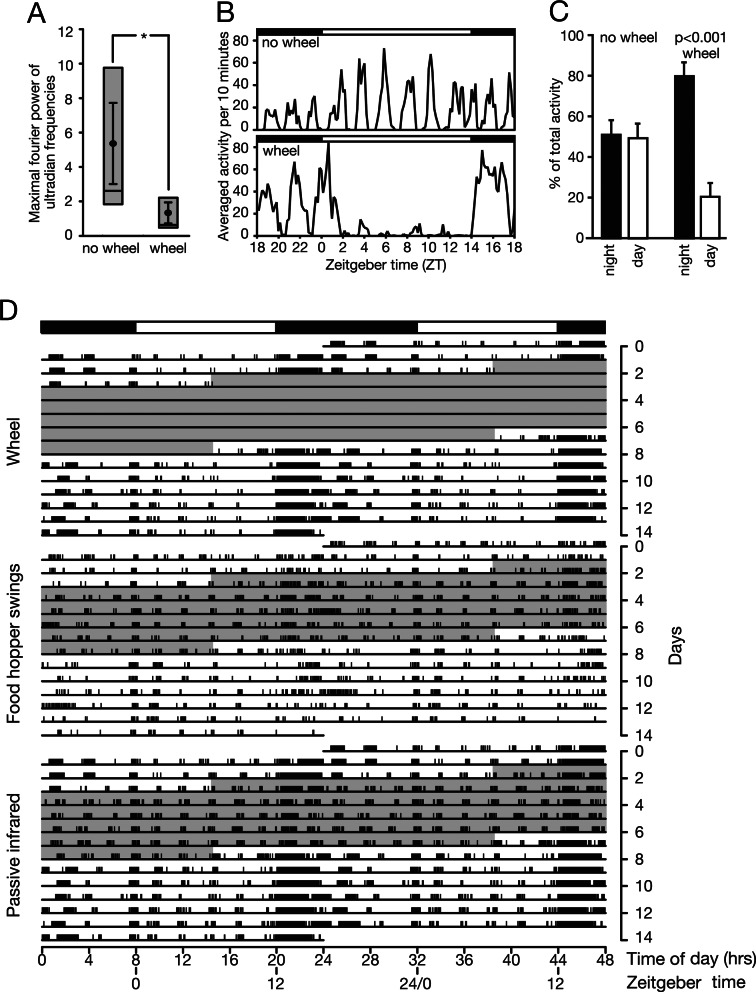
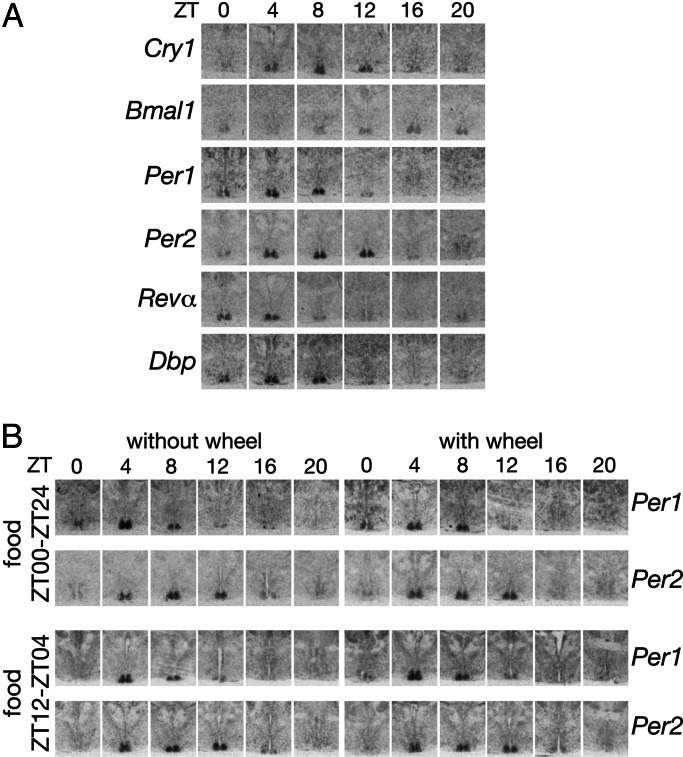
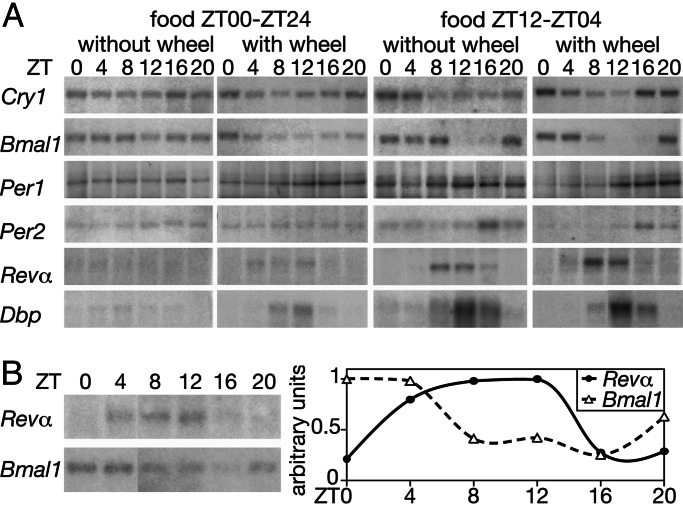
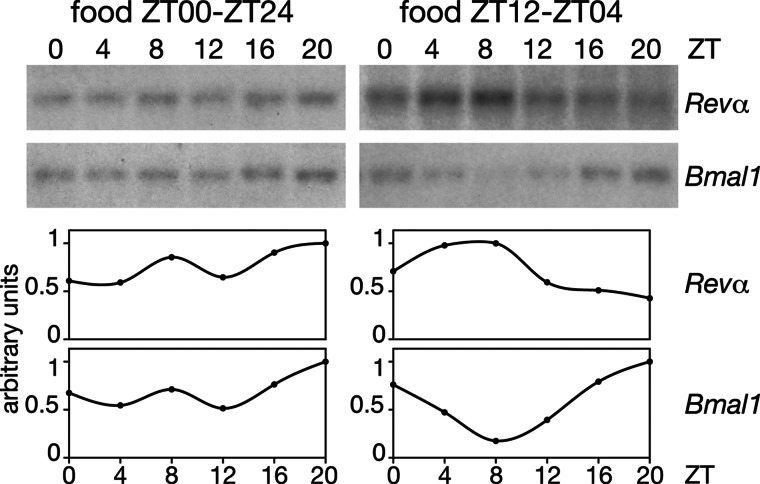
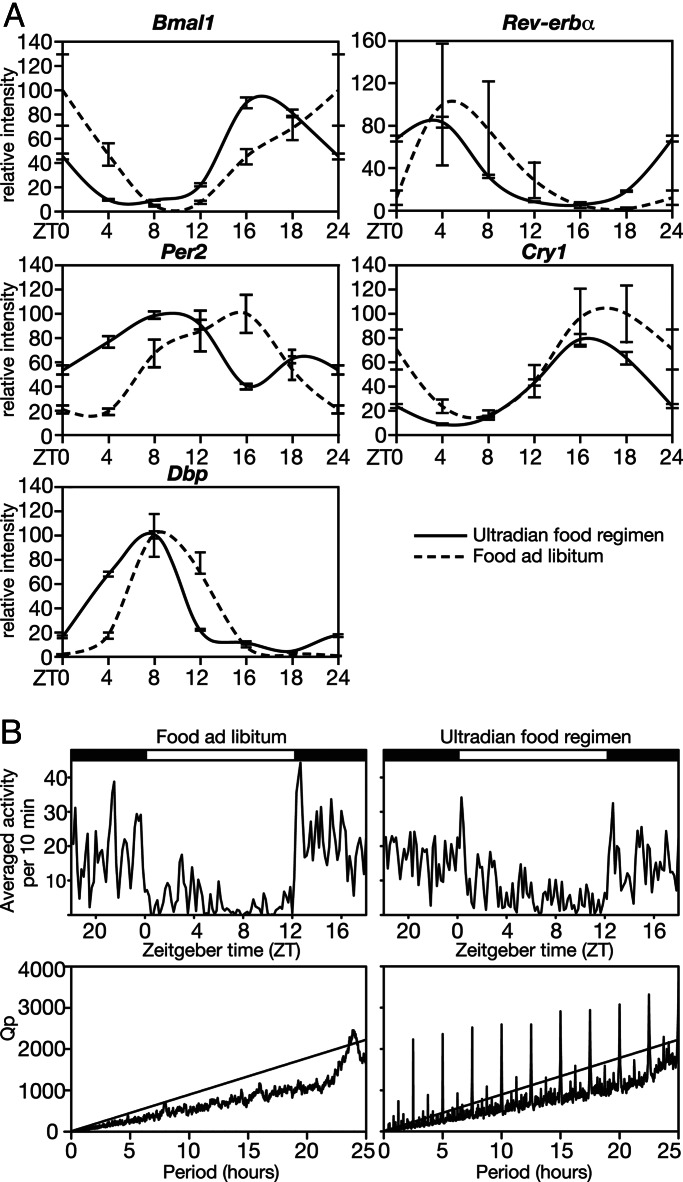
In most mammals, daily rhythms in physiology are driven by a circadian timing system composed of a master pacemaker in the suprachiasmatic nucleus (SCN) and peripheral oscillators in most body cells. The SCN clock, which is phase-entrained by light-dark cycles, is thought to synchronize subsidiary oscillators in peripheral tissues, mainly by driving cyclic feeding behavior. Here, we examined the expression of circadian clock genes in the SCN and the liver of the common vole Microtus arvalis, a rodent with ultradian activity and feeding rhythms. In these animals, clock-gene mRNAs accumulate with high circadian amplitudes in the SCN but are present at nearly constant levels in the liver. Interestingly, high-amplitude circadian liver gene expression can be elicited by subjecting voles to a circadian feeding regimen. Moreover, voles with access to a running wheel display a composite pattern of circadian and ultradian behavior, which correlates with low-amplitude circadian gene expression in the liver. Our data indicate that, in M. arvalis, the amplitude of circadian liver gene expression depends on the contribution of circadian and ultradian components in activity and feeding rhythms.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
Comment in
-
Circadian organization in the real world.Proc Natl Acad Sci U S A. 2006 Feb 28;103(9):3015-6. doi: 10.1073/pnas.0600360103. Epub 2006 Feb 21. Proc Natl Acad Sci U S A. 2006. PMID: 16492750 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
