

Modular enzyme design: regulation by mutually exclusive protein folding
- PMID: 16483603
- PMCID: PMC3145369
- DOI: 10.1016/j.jmb.2006.01.073
Modular enzyme design: regulation by mutually exclusive protein folding
Abstract
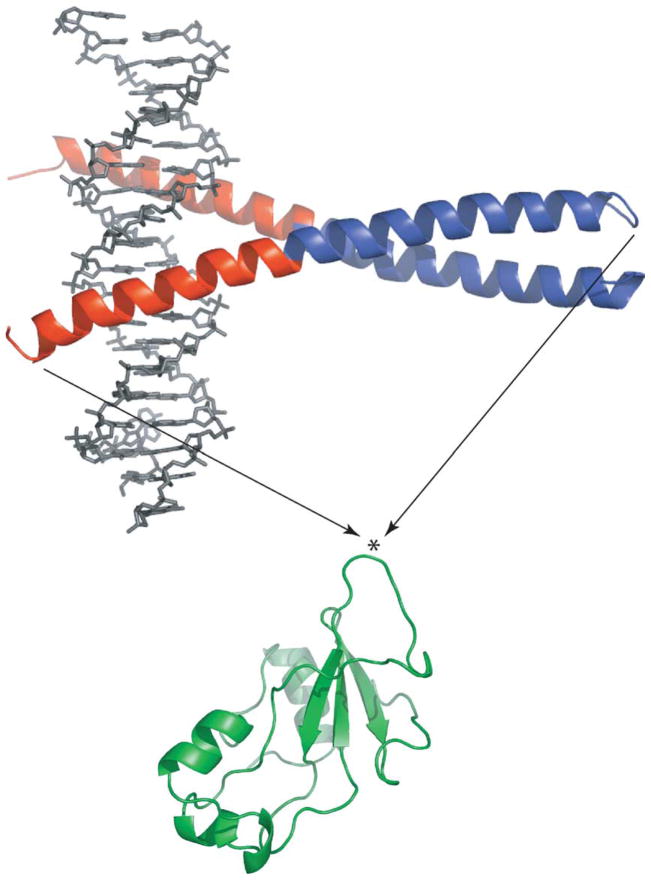
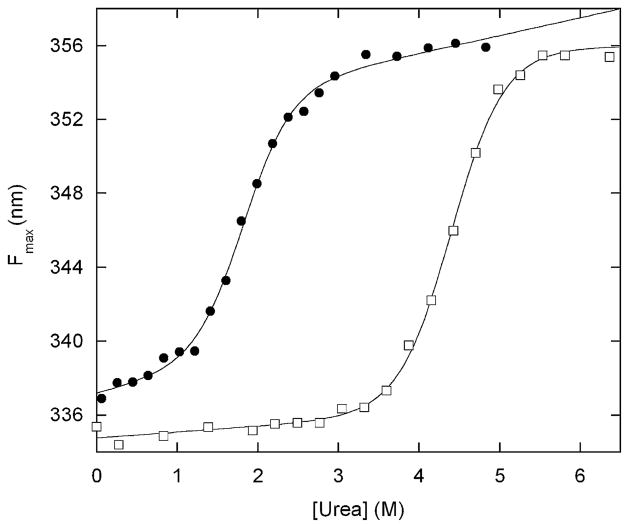
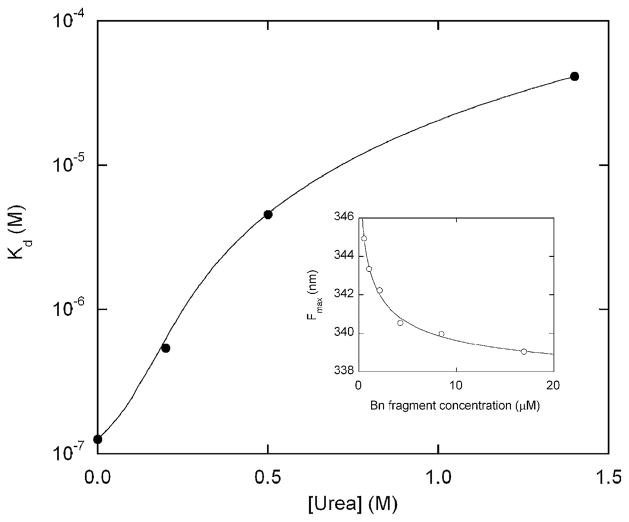
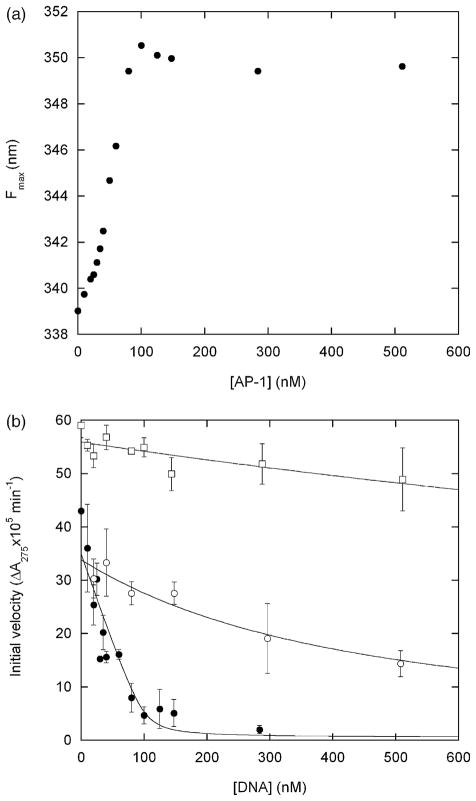
A regulatory mechanism is introduced whereupon the catalytic activity of a given enzyme is controlled by ligand binding to a receptor domain of choice. A small enzyme (barnase) and a ligand-binding polypeptide (GCN4) are fused so that a simple topological constraint prevents them from existing simultaneously in their folded states. The two domains consequently engage in a thermodynamic tug-of-war in which the more stable domain forces the less stable domain to unfold. In the absence of ligand, the barnase domain is more stable and is therefore folded and active; the GCN4 domain is substantially unstructured. DNA binding induces folding of GCN4, forcibly unfolding and inactivating the barnase domain. Barnase-GCN4 is thus a "natively unfolded" protein that uses ligand binding to switch between partially folded forms. The key characteristics of each parent protein (catalytic efficiency of barnase, DNA binding affinity and sequence specificity of GCN4) are retained in the chimera. Barnase-GCN4 thus defines a modular approach for assembling enzymes with novel sensor capabilities from a variety of catalytic and ligand binding domains.
Figures
References
-
- O’Shea EK, Klemm JD, Kim PS, Alber T. X-ray structure of the GCN4 leucine zipper, a two-stranded, parallel coiled coil. Science. 1991;254:539–544. - PubMed
-
- Cranz S, Berger C, Baici A, Jelesarov I, Bosshard HR. Monomeric and dimeric bZIP transcription factor GCN4 bind a the same rate to their target DNA site. Biochemistry. 2004;43:718–727. - PubMed
-
- Zitzewitz JA, Bilsel O, Luo J, Jones BE, Matthews CR. Probing the folding mechanism of a leucine zipper peptide by stopped-flow circular dichroism spectroscopy. Biochemistry. 1995;34:12812–12819. - PubMed
-
- Zitzewitz JA, Ibarra-Molero B, Fishel DR, Terry KL, Matthews CR. Preformed secondary structure drives the association reaction of GCN4-p1, a model coiled-coil system. J Mol Biol. 2000;286:1105–1116. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
