

Genetic variation in the Vibrio vulnificus group 1 capsular polysaccharide operon
- PMID: 16484211
- PMCID: PMC1426558
- DOI: 10.1128/JB.188.5.1987-1998.2006
Genetic variation in the Vibrio vulnificus group 1 capsular polysaccharide operon
Abstract
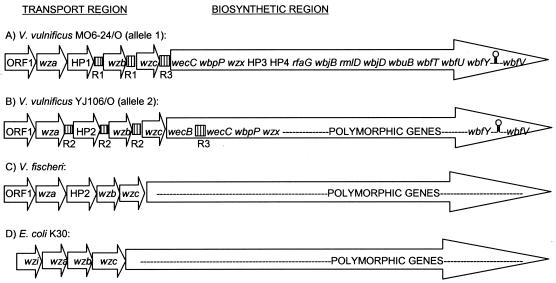
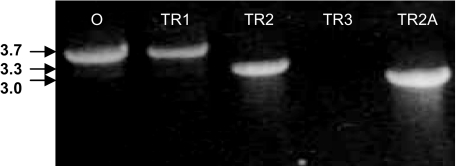
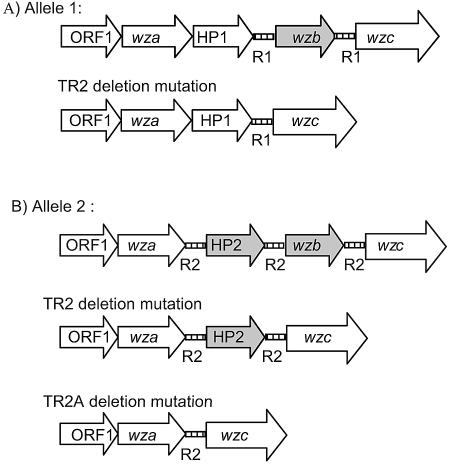
Vibrio vulnificus produces human disease associated with raw-oyster consumption or wound infections, but fatalities are limited to persons with chronic underlying illness. Capsular polysaccharide (CPS) is required for virulence, and CPS expression correlates with opaque (Op) colonies that show "phase variation" to avirulent translucent (Tr) phenotypes with reduced CPS. The results discussed here confirmed homology of a V. vulnificus CPS locus to the group 1 CPS operon in Escherichia coli. However, two distinct V. vulnificus genotypes or alleles were associated with the operon, and they diverged at sequences encoding hypothetical proteins and also at unique, intergenic repetitive DNA elements. Phase variation was examined under conditions that promoted high-frequency transition of Op to Tr forms. Recovery of Tr isolates in these experiments showed multiple genotypes, which were designated TR1, TR2, and TR3: CPS operons of TR1 isolates were identical to the Op parent, and cells remained phase variable but expressed reduced CPS. TR2 and TR3 showed deletion mutations in one (wzb) or multiple genes, respectively, and deletion mutants were acapsular and locked in the Tr phase. Complementation in trans restored the Op phenotype in strains with the wzb deletion mutation. Allelic variation in repetitive elements determined the locations, rates, and extents of deletion mutations. Thus, different mechanisms are responsible for reversible phase variation in CPS expression versus genetic deletions in the CPS operon of V. vulnificus. Repetitive-element-mediated deletion mutations were highly conserved within the species and are likely to promote survival in estuarine environments.
Figures

Similar articles
-
Identification of a group 1-like capsular polysaccharide operon for Vibrio vulnificus.Infect Immun. 2001 Nov;69(11):6893-901. doi: 10.1128/IAI.69.11.6893-6901.2001. Infect Immun. 2001. PMID: 11598064 Free PMC article.
-
Structure, Function, and Regulation of the Essential Virulence Factor Capsular Polysaccharide of Vibrio vulnificus.Int J Mol Sci. 2020 May 5;21(9):3259. doi: 10.3390/ijms21093259. Int J Mol Sci. 2020. PMID: 32380667 Free PMC article. Review.
-
Capsular polysaccharide production and serum survival of Vibrio vulnificus are dependent on antitermination control by RfaH.FEBS Lett. 2016 Dec;590(24):4564-4572. doi: 10.1002/1873-3468.12490. Epub 2016 Nov 28. FEBS Lett. 2016. PMID: 27859050
-
Evidence for an intermediate colony morphology of Vibrio vulnificus.Appl Environ Microbiol. 2006 Jun;72(6):4356-9. doi: 10.1128/AEM.02937-05. Appl Environ Microbiol. 2006. PMID: 16751551 Free PMC article.
-
Phase variation, capsular polysaccharide, pilus and flagella contribute to uptake of Vibrio vulnificus by the Eastern oyster (Crassostrea virginica).Environ Microbiol. 2009 Aug;11(8):1934-44. doi: 10.1111/j.1462-2920.2009.01916.x. Environ Microbiol. 2009. PMID: 19689704
Cited by
-
Genetic distinctions among clinical and environmental strains of Vibrio vulnificus.Appl Environ Microbiol. 2006 Sep;72(9):6136-41. doi: 10.1128/AEM.00341-06. Appl Environ Microbiol. 2006. PMID: 16957239 Free PMC article.
-
Phenotypic variation in the plant pathogenic bacterium Acidovorax citrulli.PLoS One. 2013 Sep 2;8(9):e73189. doi: 10.1371/journal.pone.0073189. eCollection 2013. PLoS One. 2013. PMID: 24023830 Free PMC article.
-
Genetic analysis and prevalence studies of the brp exopolysaccharide locus of Vibrio vulnificus.PLoS One. 2014 Jul 11;9(7):e100890. doi: 10.1371/journal.pone.0100890. eCollection 2014. PLoS One. 2014. PMID: 25013926 Free PMC article.
-
Wzb of Vibrio vulnificus represents a new group of low-molecular-weight protein tyrosine phosphatases with a unique insertion in the W-loop.J Biol Chem. 2021 Jan-Jun;296:100280. doi: 10.1016/j.jbc.2021.100280. Epub 2021 Jan 12. J Biol Chem. 2021. PMID: 33450227 Free PMC article.
-
Sticky situations: key components that control bacterial surface attachment.J Bacteriol. 2012 May;194(10):2413-25. doi: 10.1128/JB.00003-12. Epub 2012 Mar 2. J Bacteriol. 2012. PMID: 22389478 Free PMC article. Review.
References
-
- Blake, P. A., M. H. Merson, R. E. Weaver, D. G. Hollis, and P. C. Heublein. 1979. Disease caused by a marine Vibrio. Clinical characteristics and epidemiology. N. Engl. J. Med. 300:1-5. - PubMed
-
- Brosch, R., A. S. Pym, S. V. Gordon, and S. T. Cole. 2001. The evolution of mycobacterial pathogenicity: clues from comparative genomics. Trends Microbiol. 9:452-458. - PubMed
-
- Bush, C. A., P. Patel, S. Gunawardena, J. Powell, A. Joseph, J. A. Johnson, and J. G. Morris, Jr. 1997. Classification of Vibrio vulnificus strains by the carbohydrate composition of their capsular polysaccharides. Anal. Biochem. 250:186-195. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
