

Rate dependency of beta-adrenergic modulation of repolarizing currents in the guinea-pig ventricle
- PMID: 16484299
- PMCID: PMC1817790
- DOI: 10.1113/jphysiol.2006.105015
Rate dependency of beta-adrenergic modulation of repolarizing currents in the guinea-pig ventricle
Abstract
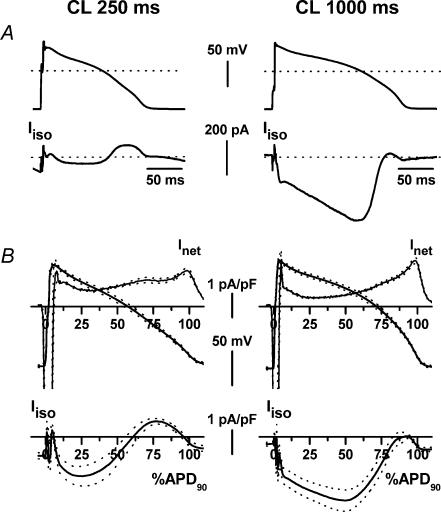
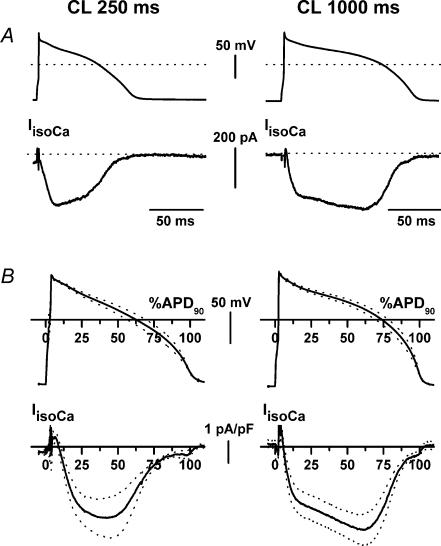
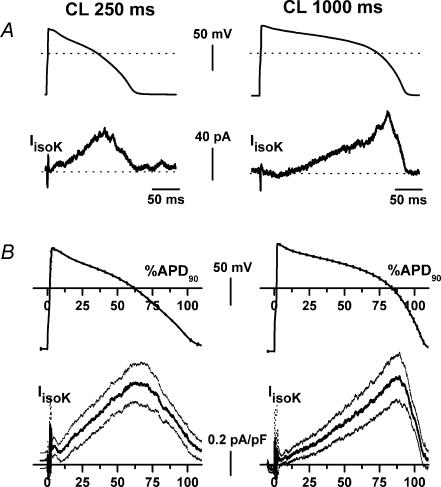
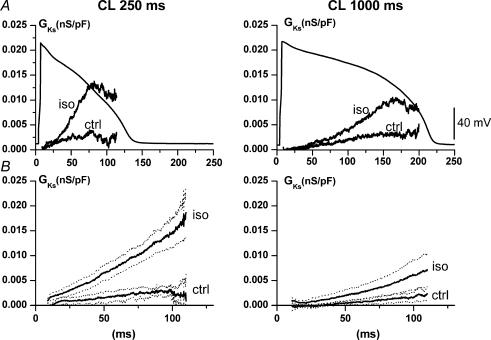
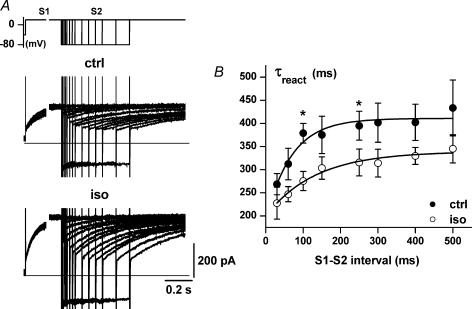
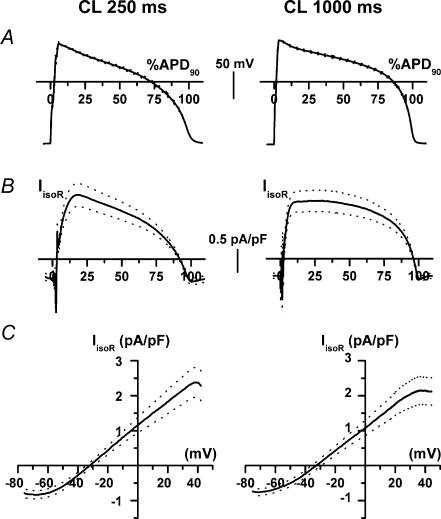
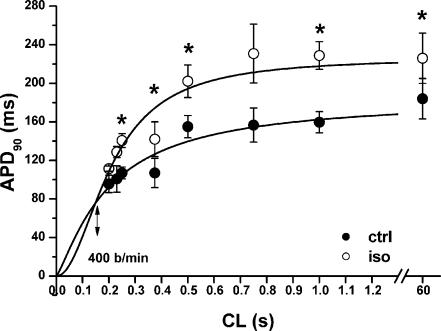
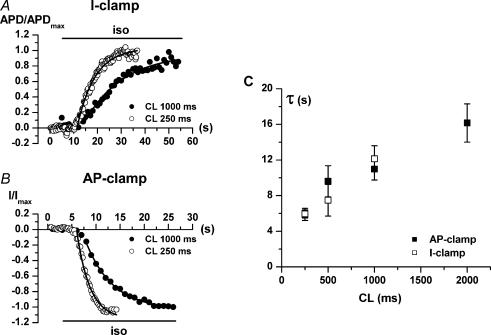
Beta-adrenergic stimulation modulates ventricular currents and sinus cycle length (CL). We investigated how changes in CL affect the current induced by isoprenaline (Iso) during the action potential (AP) of guinea-pig ventricular myocytes. Action-potential clamp was applied at CLs of 250 and 1000 ms to measure: (1) the net current induced by 0.1 microm Iso (I(Iso)); (2) the L-type Ca2+ current I(CaL) and slow delayed rectifier current I(Ks) components of I(Iso) (I(IsoCa) and I(IsoK)), identified as the Iso-induced current sensitive to nifedipine and HMR1556, respectively; and (3) I(Iso) persisting after inhibition of both I(Ca) and I(Ks) (I(isoR)). The pause dependency of I(Ks) and its modulation were evaluated in voltage-clamp experiments. The rate dependency of the duration of the action potential at 90% repolarization (APD90) and its modulation by isoprenaline were tested in current-clamp experiments. At a CL of 250 ms I(Iso) was inward during initial repolarization and reversed at 59% of APD90. At a CL of 1000 ms I(Iso) became mostly inward in all cells. Switching to shorter CL did not change I(IsoCa) and I(IsoK) amplitudes, but moved their peak amplitudes to earlier repolarization; I(IsoR) was independent of CL. Acceleration of I(IsoK) at shorter CL was based on faster pause dependency of I(Ks) activation rate. The 'restitution' of activation rates was modulated by isoprenaline. The APD90-CL relation was rotated anticlockwise by isoprenaline and crossed the control curve at a CL of 150 ms (400 beats min(-1)). We conclude that: (1) isoprenaline induced markedly different current profiles according to pacing rate, involving CL-dependent I(Ca) and I(Ks) modulation; (2) the effect of isoprenaline on APD90 was CL dependent, and negligible during tachycardia; and (3) during sympathetic activation, repolarization stability may involve matched modulation of sinus rate and repolarizing currents.
Figures
Similar articles
-
Mechanisms of beta-adrenergic modulation of I(Ks) in the guinea-pig ventricle: insights from experimental and model-based analysis.Biophys J. 2009 May 6;96(9):3862-72. doi: 10.1016/j.bpj.2009.02.017. Biophys J. 2009. PMID: 19413992 Free PMC article.
-
I(Ks) restricts excessive beat-to-beat variability of repolarization during beta-adrenergic receptor stimulation.J Mol Cell Cardiol. 2010 Jan;48(1):122-30. doi: 10.1016/j.yjmcc.2009.08.033. Epub 2009 Sep 8. J Mol Cell Cardiol. 2010. PMID: 19744496
-
Beta-Adrenergic Activation of the Inward Rectifier K+ Current Is Mediated by the CaMKII Pathway in Canine Ventricular Cardiomyocytes.Int J Mol Sci. 2024 Oct 29;25(21):11609. doi: 10.3390/ijms252111609. Int J Mol Sci. 2024. PMID: 39519160 Free PMC article.
-
Anti-adrenergic effect of adenosine on Na(+)-Ca(2+) exchange current recorded from guinea-pig ventricular myocytes.Cell Calcium. 2001 May;29(5):347-58. doi: 10.1054/ceca.2001.0197. Cell Calcium. 2001. PMID: 11292391
-
Rate dependency of delayed rectifier currents during the guinea-pig ventricular action potential.J Physiol. 2001 Aug 1;534(Pt 3):721-32. doi: 10.1111/j.1469-7793.2001.00721.x. J Physiol. 2001. PMID: 11483703 Free PMC article.
Cited by
-
Altered K+ current profiles underlie cardiac action potential shortening in hyperkalemia and β-adrenergic stimulation.Can J Physiol Pharmacol. 2019 Aug;97(8):773-780. doi: 10.1139/cjpp-2019-0056. Epub 2019 May 15. Can J Physiol Pharmacol. 2019. PMID: 31091413 Free PMC article.
-
Calsequestrin mutation and catecholaminergic polymorphic ventricular tachycardia: a simulation study of cellular mechanism.Cardiovasc Res. 2007 Jul 1;75(1):79-88. doi: 10.1016/j.cardiores.2007.04.010. Epub 2007 Apr 21. Cardiovasc Res. 2007. PMID: 17531962 Free PMC article.
-
Dynamic Action Potential Restitution Contributes to Mechanical Restitution in Right Ventricular Myocytes From Pulmonary Hypertensive Rats.Front Physiol. 2018 Mar 12;9:205. doi: 10.3389/fphys.2018.00205. eCollection 2018. Front Physiol. 2018. PMID: 29593564 Free PMC article.
-
Differential roles of two delayed rectifier potassium currents in regulation of ventricular action potential duration and arrhythmia susceptibility.J Physiol. 2017 Apr 1;595(7):2301-2317. doi: 10.1113/JP273191. Epub 2016 Dec 28. J Physiol. 2017. PMID: 27779762 Free PMC article.
-
Exploring the Coordination of Cardiac Ion Channels With Action Potential Clamp Technique.Front Physiol. 2022 Mar 16;13:864002. doi: 10.3389/fphys.2022.864002. eCollection 2022. Front Physiol. 2022. PMID: 35370800 Free PMC article. Review.
References
-
- Doerr T, Denger R, Doerr A, Trautwein W. Ionic currents contributing to the action potential in single ventricular myocytes of the guinea pig studied with action potential clamp. Pflugers Arch. 1990;416:230–237. - PubMed
-
- Fabiato A, Fabiato F. Calculator programs for computing the composition of the solutions containing multiple metals and ligands used for experiments in skinned muscle cells. J Physiol (Paris) 1979;75:463–505. - PubMed
-
- Gladman G, Davis AM, Fogelman R, Hamilton RM, Gow RM. Torsade de pointes, acquired complete heart block and inappropriately long QT in childhood. Can J Cardiol. 1996;12:683–685. - PubMed
-
- Hamlin RL, Kijtawornrat A, Keene BW, Hamlin DM. QT and RR intervals in conscious and anesthetized guinea pigs with highly varying RR intervals and given QTc-lengthening test articles. Toxicol Sci. 2003;76:437–442. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous