

Phosphatidylinositol-3-phosphate kinase pathway activation protects leukemic large granular lymphocytes from undergoing homeostatic apoptosis
- PMID: 16484592
- PMCID: PMC1895814
- DOI: 10.1182/blood-2005-08-3076
Phosphatidylinositol-3-phosphate kinase pathway activation protects leukemic large granular lymphocytes from undergoing homeostatic apoptosis
Abstract
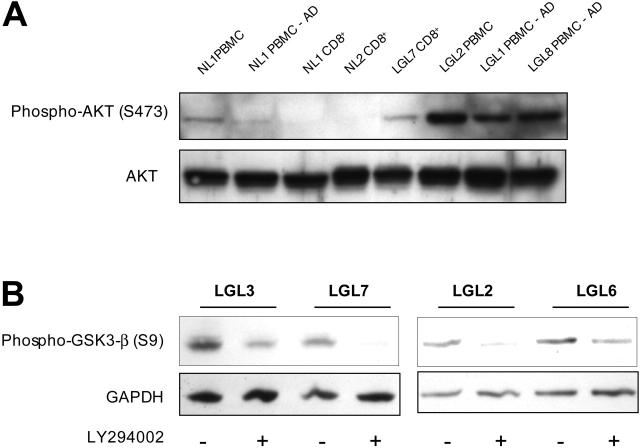
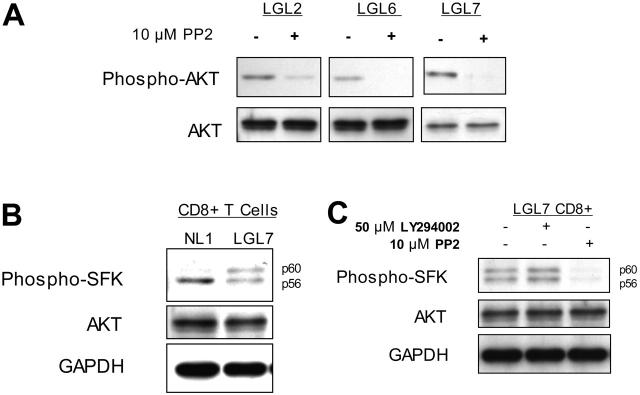
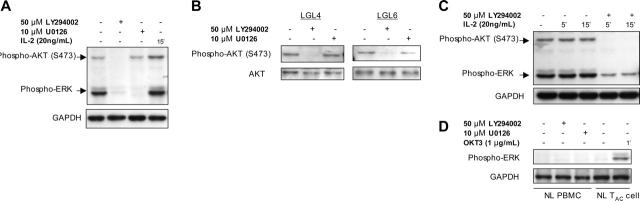
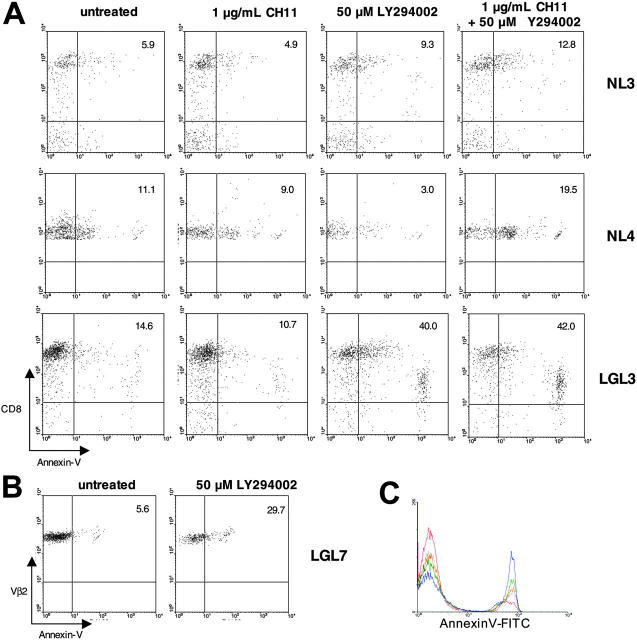
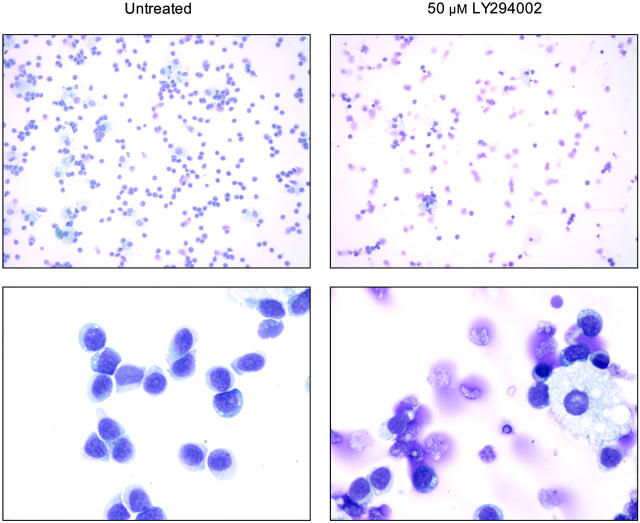
T-cell large granular lymphocytic leukemia (T-LGL) is characterized by chronic clonal lymphoproliferation of cytotoxic T lymphocytes (CTLs). Despite exhibiting phenotypic properties of antigen-activated cells, including expression of Fas and FasL, T-LGL cells accumulate and demonstrate resistance to apoptosis. We propose that increased activity of a prosurvival signaling pathway in T-LGL is responsible for attenuated apoptosis in T-LGL. Given the importance of the phosphatidylinositol-3 kinase (PI3K)-AKT pathway in regulating the balance between survival and apoptosis, we analyzed AKT activity in T-LGL cells. Compared with resting CTLs from healthy donors, patients' T-LGL cells showed higher levels of phosphorylated AKT. We demonstrate that phospho-AKT induction is dependent on the upstream activity of a Src family kinase. Since the PI3K-AKT pathway can antagonize the ability of Fas to initiate apoptosis, we hypothesized that inhibition of PI3K would lead to reacquisition of Fas sensitivity in T-LGL. Inhibition of the PI3K-AKT pathway alone led to brisk spontaneous apoptosis of T-LGL. These results suggest that T-LGL pathogenesis is dependent on activity of the PI3K-AKT pathway, without which the leukemic cells will begin to undergo spontaneous apoptosis. We propose that novel therapeutics inhibiting the PI3K-AKT axis may provide effective treatment for T-LGL.
Figures
References
-
- Melenhorst JJ, Sorbara L, Kirby M, Hensel NF, Barrett AJ. Large granular lymphocyte leukaemia is characterized by a clonal T-cell receptor rearrangement in both memory and effector CD8(+) lymphocyte populations. Br J Haematol. 2001;112: 189-194. - PubMed
-
- Lamy T, Liu JH, Landowski TH, Dalton WS, Loughran TP Jr. Dysregulation of CD95/CD95 ligand-apoptotic pathway in CD3(+) large granular lymphocyte leukemia. Blood. 1998;92: 4771-4777. - PubMed
-
- Kothapalli R, Nyland SB, Kusmartseva I, Bailey RD, McKeown TM, Loughran TP Jr. Constitutive production of proinflammatory cytokines RAN-TES, MIP-1beta and IL-18 characterizes LGL leukemia. Int J Oncol. 2005;26: 529-535. - PubMed
-
- Liu JH, Wei S, Lamy T, et al. Chronic neutropenia mediated by fas ligand. Blood. 2000;95: 3219-3222. - PubMed
-
- Shvidel L, Duksin C, Tzimanis A, et al. Cytokine release by activated T-cells in large granular lymphocytic leukemia associated with autoimmune disorders. Hematol J. 2002;3: 32-37. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
