

Processing stages underlying word recognition in the anteroventral temporal lobe
- PMID: 16488158
- PMCID: PMC1513618
- DOI: 10.1016/j.neuroimage.2005.10.053
Processing stages underlying word recognition in the anteroventral temporal lobe
Abstract
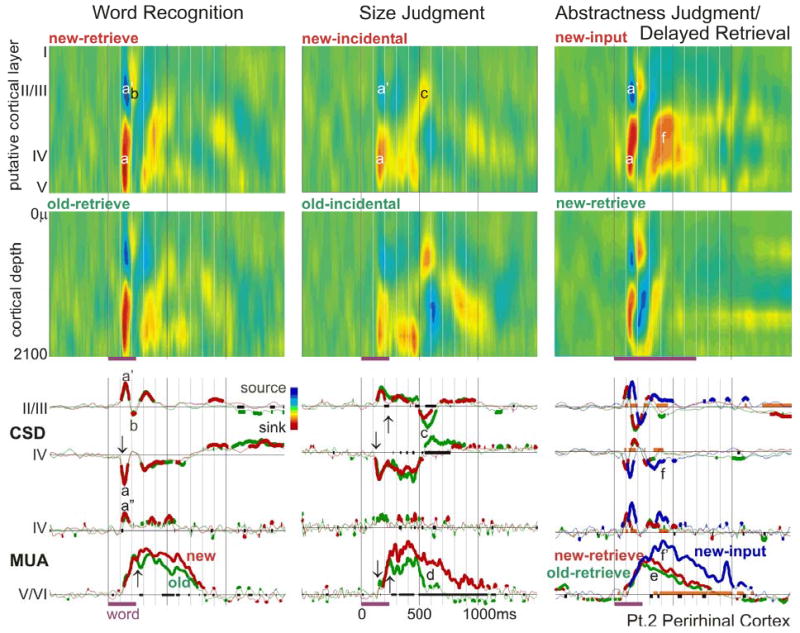
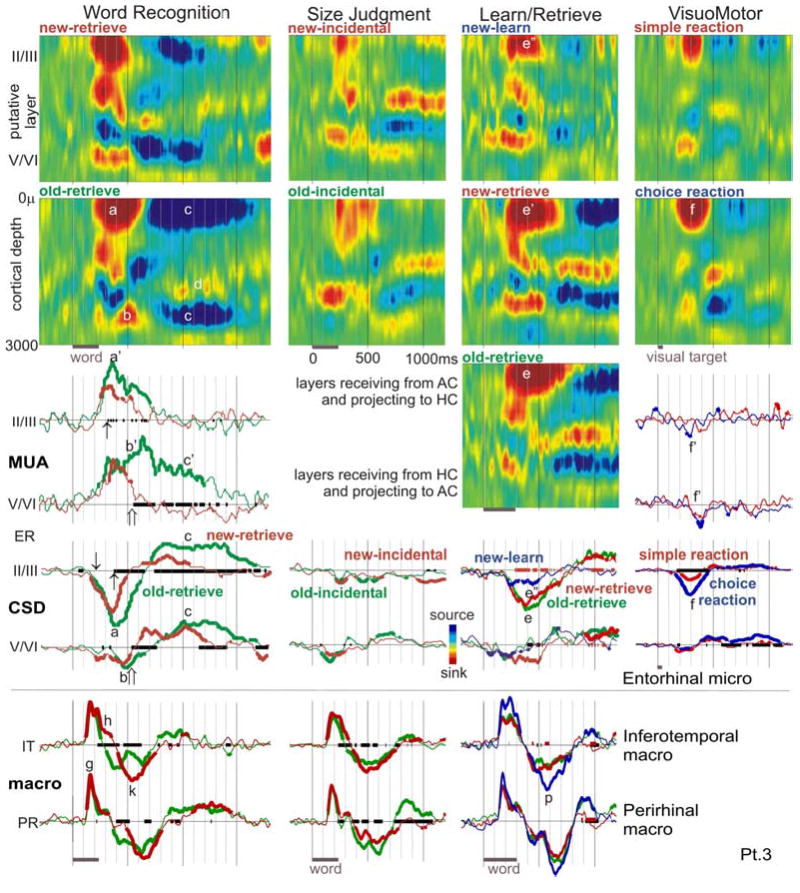
The anteroventral temporal lobe integrates visual, lexical, semantic and mnestic aspects of word processing, through its reciprocal connections with the ventral visual stream, language areas, and the hippocampal formation. We used linear microelectrode arrays to probe population synaptic currents and neuronal firing in different cortical layers of the anteroventral temporal lobe, during semantic judgments with implicit priming and overt word recognition. Since different extrinsic and associative inputs preferentially target different cortical layers, this method can help reveal the sequence and nature of local processing stages at a higher resolution than was previously possible. The initial response in inferotemporal and perirhinal cortices is a brief current sink beginning at approximately 120 ms and peaking at approximately 170 ms. Localization of this initial sink to middle layers suggests that it represents feedforward input from lower visual areas, and simultaneously increased firing implies that it represents excitatory synaptic currents. Until approximately 800 ms, the main focus of transmembrane current sinks alternates between middle and superficial layers, with the superficial focus becoming increasingly dominant after approximately 550 ms. Since superficial layers are the target of local and feedback associative inputs, this suggests an alternation in predominant synaptic input between feedforward and feedback modes. Word repetition does not affect the initial perirhinal and inferotemporal middle layer sink but does decrease later activity. Entorhinal activity begins later (approximately 200 ms), with greater apparent excitatory post-synaptic currents and multiunit activity in neocortically projecting than hippocampal-projecting layers. In contrast to perirhinal and entorhinal responses, entorhinal responses are larger to repeated words during memory retrieval. These results identify a sequence of physiological activation, beginning with a sharp activation from lower level visual areas carrying specific information to middle layers. This is followed by feedback and associative interactions involving upper cortical layers, which are abbreviated to repeated words. Following bottom-up and associative stages, top-down recollective processes may be driven by entorhinal cortex. Word processing involves a systematic sequence of fast feedforward information transfer from visual areas to anteroventral temporal cortex followed by prolonged interactions of this feedforward information with local associations and feedback mnestic information from the medial temporal lobe.
Figures


References
-
- Allison T, Puce A, Spencer DD, McCarthy G. Electrophysiological studies of human face perception. I: Potentials generated in occipitotemporal cortex by face and non-face stimuli. Cereb Cortex. 1999;9:415–430. - PubMed
-
- Bancaud J, Brunet-Bourgin F, Chauvel P, Halgren E. Anatomical origin of déjà vu and vivid 'memories' in human temporal lobe epilepsy. Brain. 1994;117:71–90. - PubMed
-
- Barbas H, Rempel-Clower N. Cortical structure predicts the pattern of corticocortical connections. Cereb Cortex. 1997;7:635–646. - PubMed
-
- Barth DS, Di S. Three-dimensional analysis of auditory-evoked potentials in rat neocortex. J Neurophysiol. 1990;64:1527–1536. - PubMed
-
- Bartolomei F, Barbeau E, Gavaret M, Guye M, McGonigal A, Regis J, Chauvel P. Cortical stimulation study of the role of rhinal cortex in deja vu and reminiscence of memories. Neurology. 2004;63:858–864. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
