

Exploring the ESCRTing machinery in eukaryotes
- PMID: 16488176
- PMCID: PMC2865992
- DOI: 10.1016/j.tplants.2006.01.008
Exploring the ESCRTing machinery in eukaryotes
Abstract
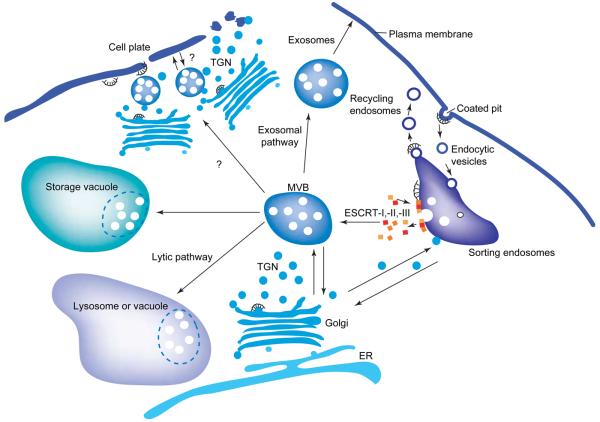
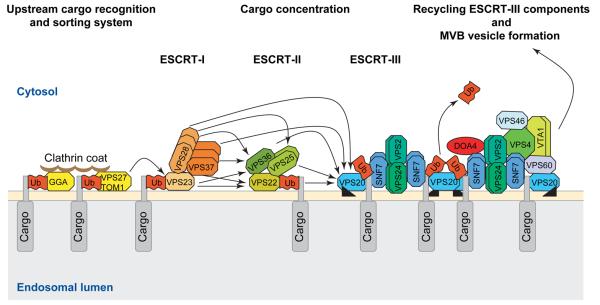
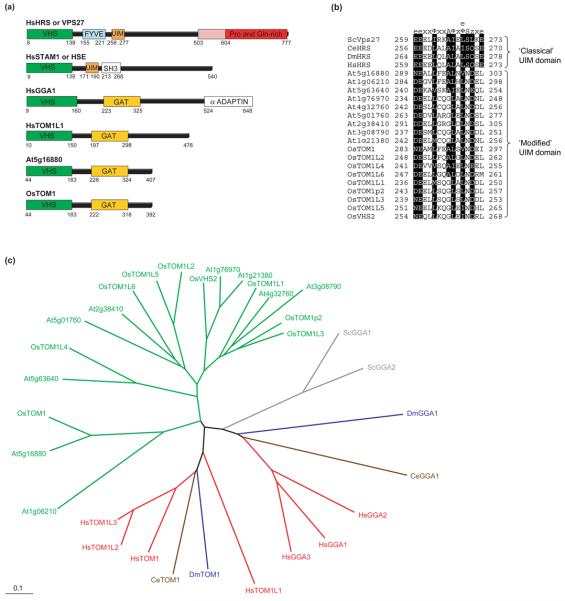
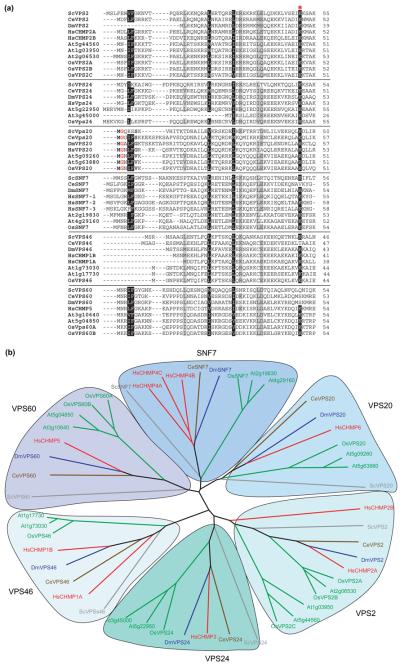
The profile of protein sorting into multivesicular bodies (MVBs) has risen recently with the identification of three heteromeric complexes known as ESCRT-I,-II,-III (Endosomal Sorting Complex Required for Transport). Genetic analyses in yeast have identified up to 15 soluble class E VPS (vacuolar protein sorting) proteins that have been assigned to the ESCRT machinery and function in cargo recognition and sorting, complex assembly, vesicle formation and dissociation. Despite their functional importance in yeast and mammalian cells, little is known about their presence and function in other organisms including plants. We have made use of the fully sequenced genomes of Arabidopsis thaliana and Oryza sativa, Drosophila melanogaster and Caenorhabditis elegans to explore the identity, structural characteristics and phylogenetic relationships of proteins assigned to the ESCRT machinery.
Figures
References
-
- Tanchak MA, Fowke LC. The morphology of multivesicular bodies in soybean protoplasts and their role in endocytosis. Protoplasma. 1987;138:173–182.
-
- Marcote MJ, et al. Membrane transport in the endocytic pathway: animal versus plant cells. Protoplasma. 2000;210:123–132.
-
- Jiang L, et al. Multivesicular bodies: a mechanism to package lytic and storage functions in one organelle? Trends Cell Biol. 2002;12:362–367. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
