

Novel thermostable Y-family polymerases: applications for the PCR amplification of damaged or ancient DNAs
- PMID: 16488882
- PMCID: PMC1373694
- DOI: 10.1093/nar/gkj512
Novel thermostable Y-family polymerases: applications for the PCR amplification of damaged or ancient DNAs
Abstract
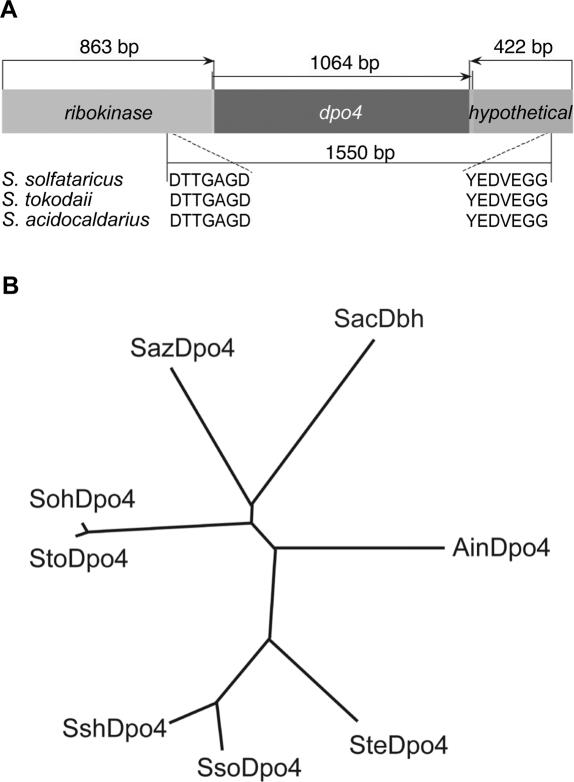
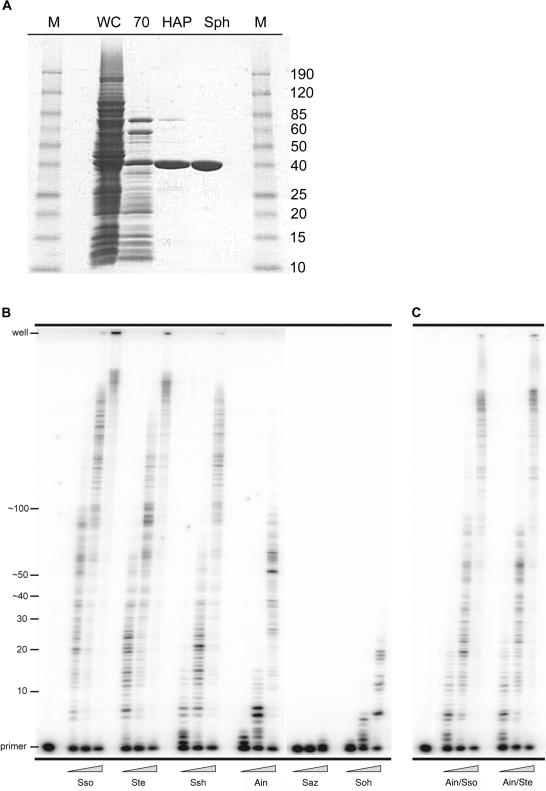
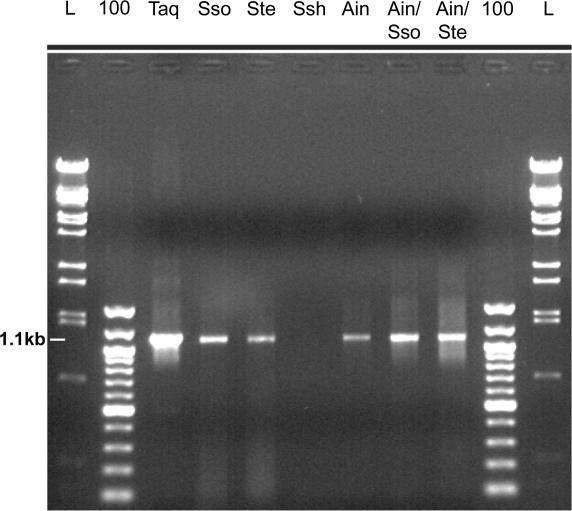
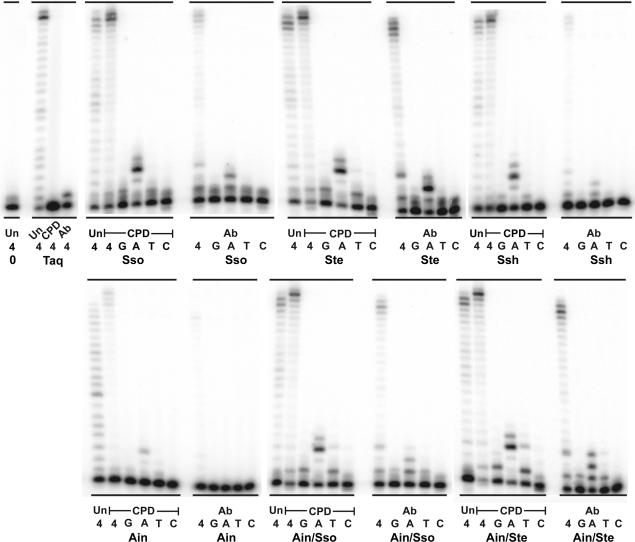
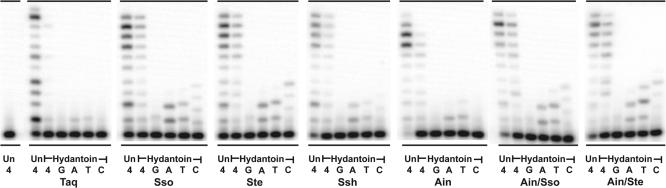
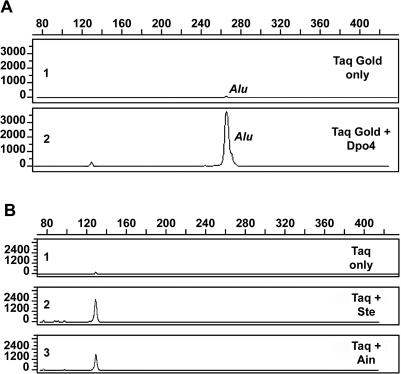
For many years, Taq polymerase has served as the stalwart enzyme in the PCR amplification of DNA. However, a major limitation of Taq is its inability to amplify damaged DNA, thereby restricting its usefulness in forensic applications. In contrast, Y-family DNA polymerases, such as Dpo4 from Sulfolobus solfataricus, can traverse a wide variety of DNA lesions. Here, we report the identification and characterization of five novel thermostable Dpo4-like enzymes from Acidianus infernus, Sulfolobus shibatae, Sulfolobus tengchongensis, Stygiolobus azoricus and Sulfurisphaera ohwakuensis, as well as two recombinant chimeras that have enhanced enzymatic properties compared with the naturally occurring polymerases. The Dpo4-like polymerases are moderately processive, can substitute for Taq in PCR and can bypass DNA lesions that normally block Taq. Such properties make the Dpo4-like enzymes ideally suited for the PCR amplification of damaged DNA samples. Indeed, by using a blend of Taq and Dpo4-like enzymes, we obtained a PCR amplicon from ultraviolet-irradiated DNA that was largely unamplifyable with Taq alone. The inclusion of thermostable Dpo4-like polymerases in PCRs, therefore, augments the recovery and analysis of lesion-containing DNA samples, such as those commonly found in forensic or ancient DNA molecular applications.
Figures
References
-
- Ohmori H., Friedberg E.C., Fuchs R.P.P., Goodman M.F., Hanaoka F., Hinkle D., Kunkel T.A., Lawrence C.W., Livneh Z., Nohmi T., et al. The Y-family of DNA polymerases. Mol. Cell. 2001;8:7–8. - PubMed
-
- Kulaeva O.I., Koonin E.V., McDonald J.P., Randall S.K., Rabinovich N., Connaughton J.F., Levine A.S., Woodgate R. Identification of a DinB/UmuC homolog in the archeon Sulfolobus solfataricus. Mutat. Res. 1996;357:245–253. - PubMed
-
- Gruz P., Pisani F.M., Shimizu M., Yamada M., Hayashi I., Morikawa K., Nohmi T. Synthetic activity of Sso DNA polymerase Y1, an archaeal DinB-like DNA polymerase, is stimulated by processivity factors proliferating cell nuclear antigen and replication factor C. J. Biol. Chem. 2001;276:47394–47401. - PubMed
-
- Potapova O., Grindley N.D., Joyce C.M. The mutational specificity of the dbh lesion bypass polymerase and its implications. J. Biol. Chem. 2002;277:28157–28166. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
