

A simple and robust statistical test for detecting the presence of recombination
- PMID: 16489234
- PMCID: PMC1456386
- DOI: 10.1534/genetics.105.048975
A simple and robust statistical test for detecting the presence of recombination
Abstract
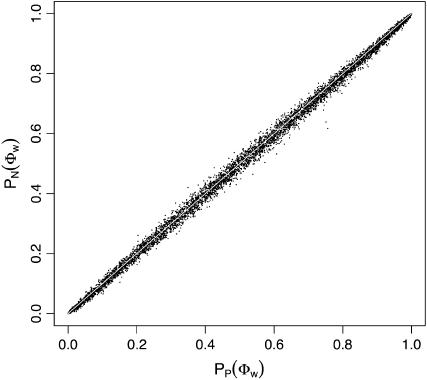
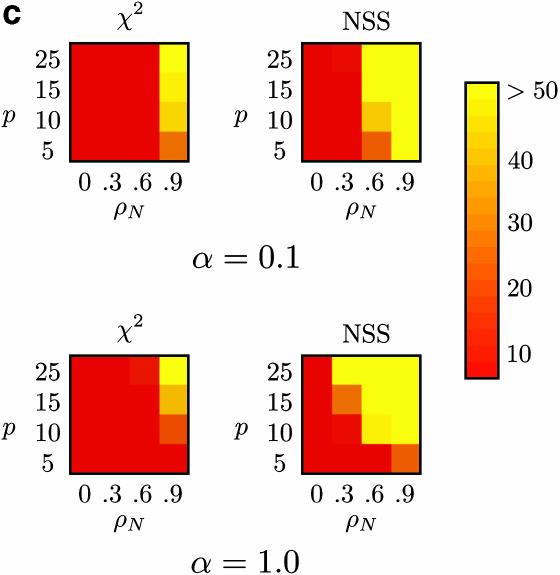
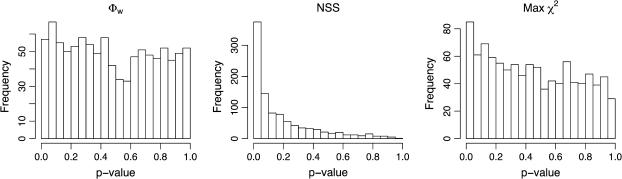
Recombination is a powerful evolutionary force that merges historically distinct genotypes. But the extent of recombination within many organisms is unknown, and even determining its presence within a set of homologous sequences is a difficult question. Here we develop a new statistic, phi(w), that can be used to test for recombination. We show through simulation that our test can discriminate effectively between the presence and absence of recombination, even in diverse situations such as exponential growth (star-like topologies) and patterns of substitution rate correlation. A number of other tests, Max chi2, NSS, a coalescent-based likelihood permutation test (from LDHat), and correlation of linkage disequilibrium (both r2 and /D'/) with distance, all tend to underestimate the presence of recombination under strong population growth. Moreover, both Max chi2 and NSS falsely infer the presence of recombination under a simple model of mutation rate correlation. Results on empirical data show that our test can be used to detect recombination between closely as well as distantly related samples, regardless of the suspected rate of recombination. The results suggest that phi(w) is one of the best approaches to distinguish recurrent mutation from recombination in a wide variety of circumstances.
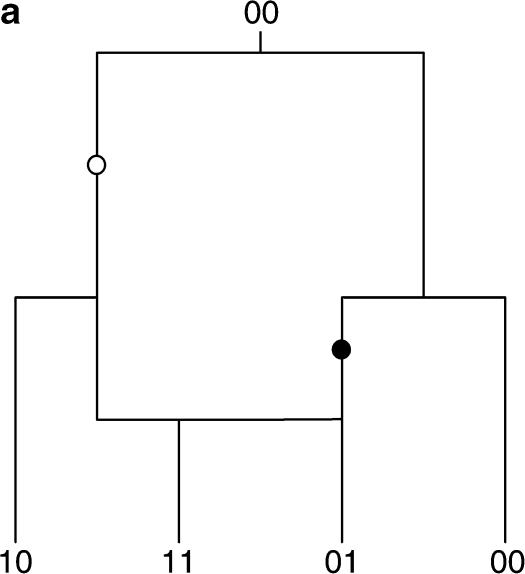
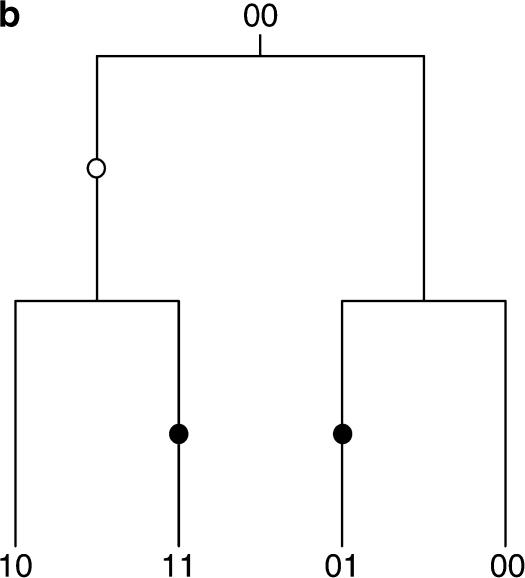
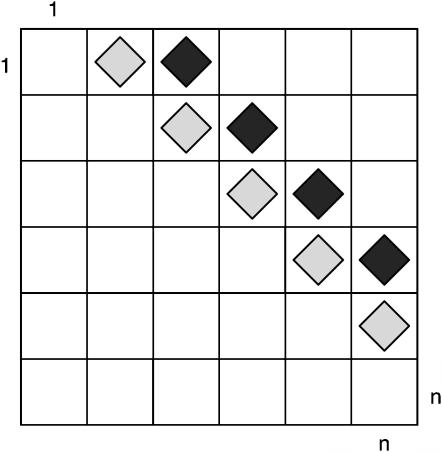
Figures
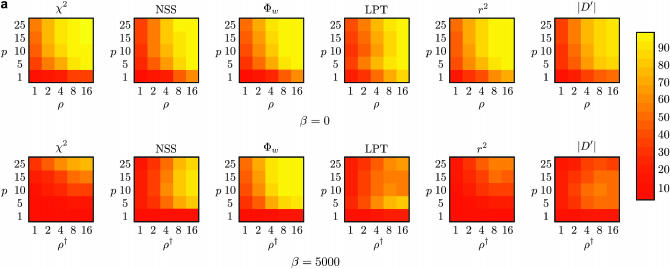
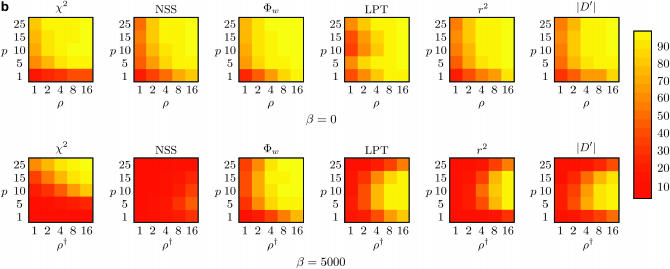
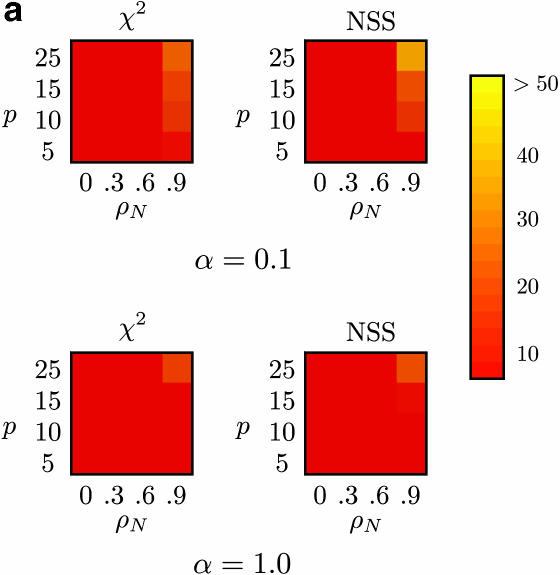
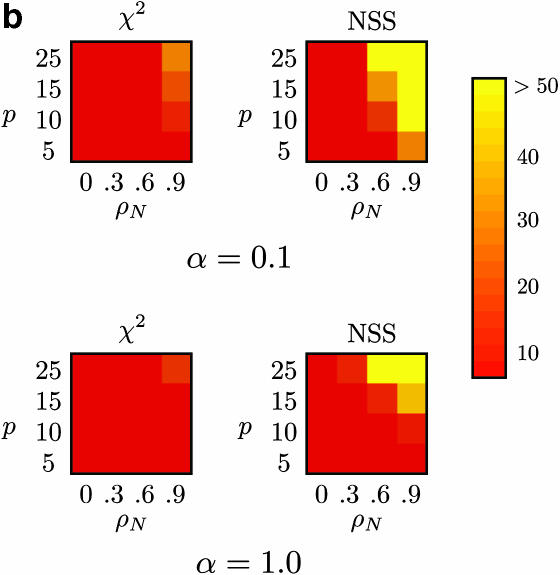
References
-
- Awadalla, P., 2003. The evolutionary genomics of pathogen recombination. Nat. Rev. Genet. 4(1): 50–60. - PubMed
-
- Awadalla, P., A. Eyre-Walker and J. M. Smith, 1999. Linkage disequilibrium and recombination in hominid mitochondrial DNA. Science 286(5449): 2524–2525. - PubMed
-
- Brown, C. J., E. C. Garner, A. Keith Dunker and P. Joyce, 2001. The power to detect recombination using the coalescent. Mol. Biol. Evol. 18(7): 1421–1424. - PubMed
-
- Bruen, T., and D. Bryant, 2006. A subdivision approach to maximum parsimony. Ann. Combinator. (in press).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
