

Rhodopsin self-associates in asolectin liposomes
- PMID: 16492772
- PMCID: PMC1413906
- DOI: 10.1073/pnas.0511010103
Rhodopsin self-associates in asolectin liposomes
Abstract
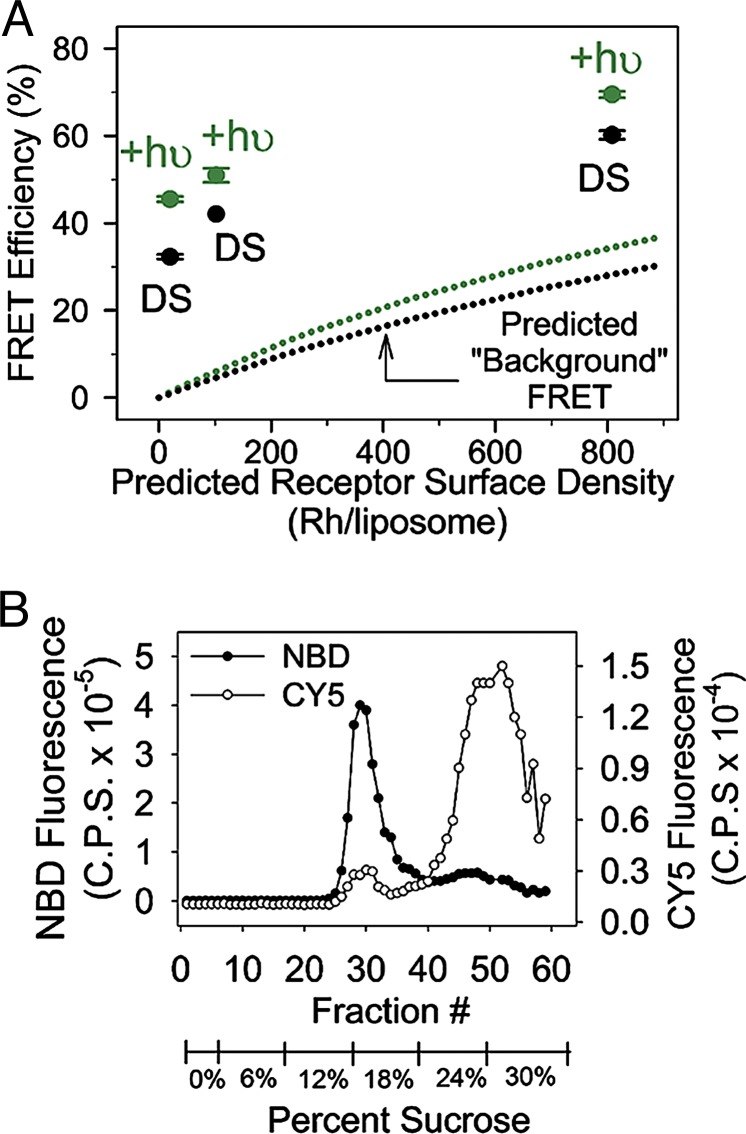
We show that the photoreceptor rhodopsin (Rh) can exist in the membrane as a dimer or multimer using luminescence resonance energy transfer and FRET methods. Our approach looked for interactions between Rh molecules reconstituted into asolectin liposomes. The low receptor density used in the measurements ensured minimal receptor crowding and artifactual association. The fluorescently labeled Rh molecules were fully functional, as measured by their ability to activate the G protein transducin. The luminescence resonance energy transfer measurements revealed a distance of 47-50 Angstroms between Rh molecules. The measured efficiency of FRET between receptors was close to the theoretical maximum possible, indicating nearly quantitative Rh-Rh association. Together, these results provide compelling evidence that Rh spontaneously self-associates in membranes.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
