

High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize
- PMID: 16492777
- PMCID: PMC1413895
- DOI: 10.1073/pnas.0509650103
High frequency of centromere inactivation resulting in stable dicentric chromosomes of maize
Abstract
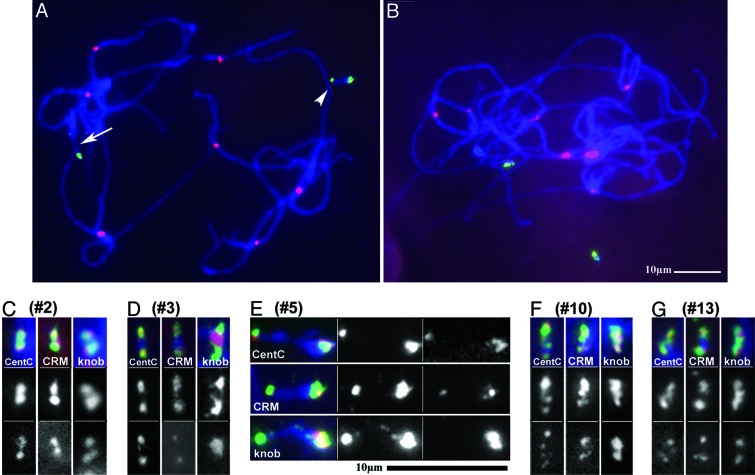
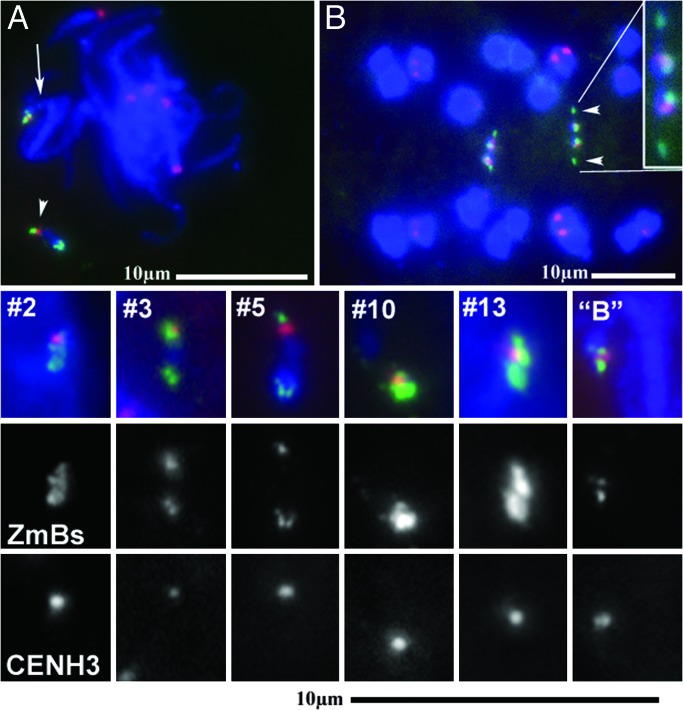
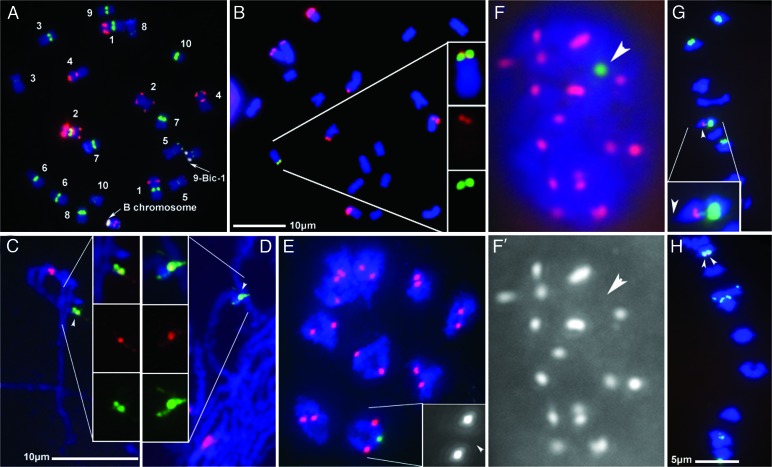
Somatic chromosome spreads from maize (Zea mays L.) plants containing B-A translocation chromosomes undergoing the chromosome type breakage-fusion-bridge cycle were examined by FISH. The size and type of extra chromosomes varied among cells of the same individual. A collection of minichromosomes derived from the chromosome type breakage-fusion-bridge cycle was examined for the presence of stable dicentric chromosomes. Six of 23 chromosomes in the collection contained two regions with DNA sequences typical of centromeres. Functional analysis and immunolabeling of CENH3, the centromere-specific histone H3 variant, revealed only one functional centromere per chromosome, despite the duplicate centromere sequences. One plant was found with an inactive B centromere that had been translocated to the short arm of chromosome 9. The translocated centromere region appeared identical to that of a normal B chromosome. The inactivation of the centromeres was stable for at least four generations. By using dicentrics from dispensable chromosomes, centromere inactivation was found to be quite common under these circumstances.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures


References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
