

Directly reactivated, but not indirectly reactivated, memories undergo reconsolidation in the amygdala
- PMID: 16492789
- PMCID: PMC1413871
- DOI: 10.1073/pnas.0507168103
Directly reactivated, but not indirectly reactivated, memories undergo reconsolidation in the amygdala
Abstract
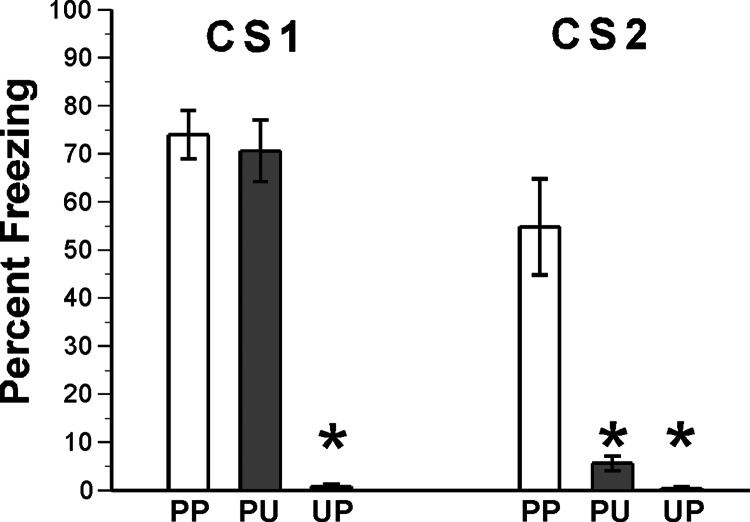
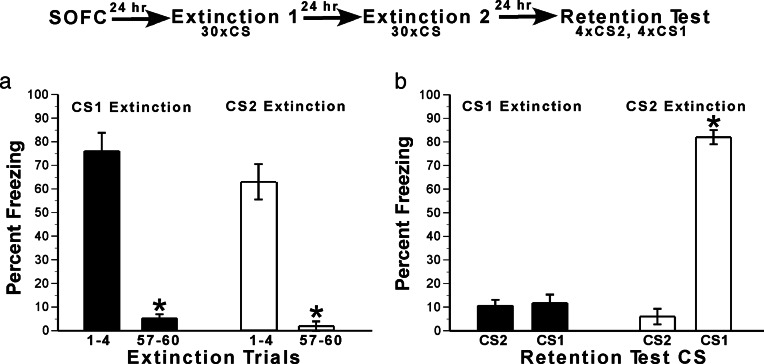
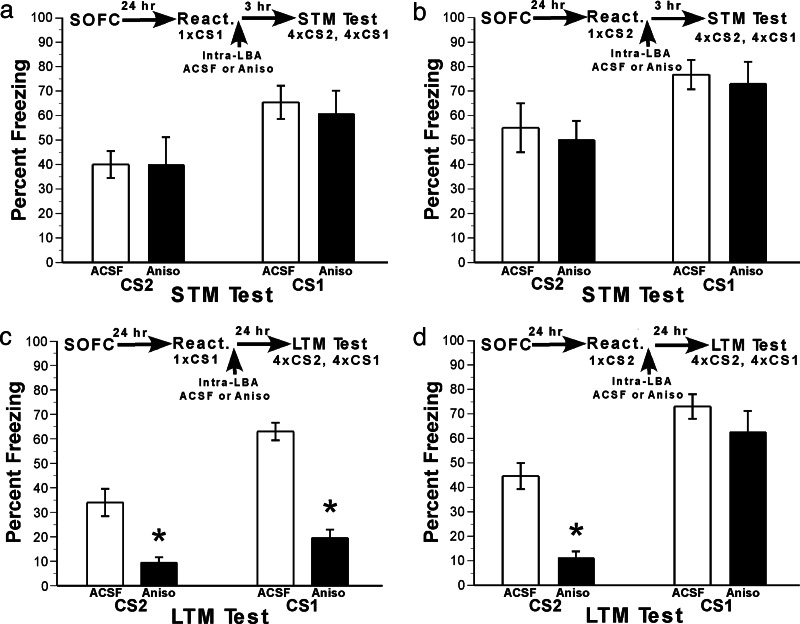
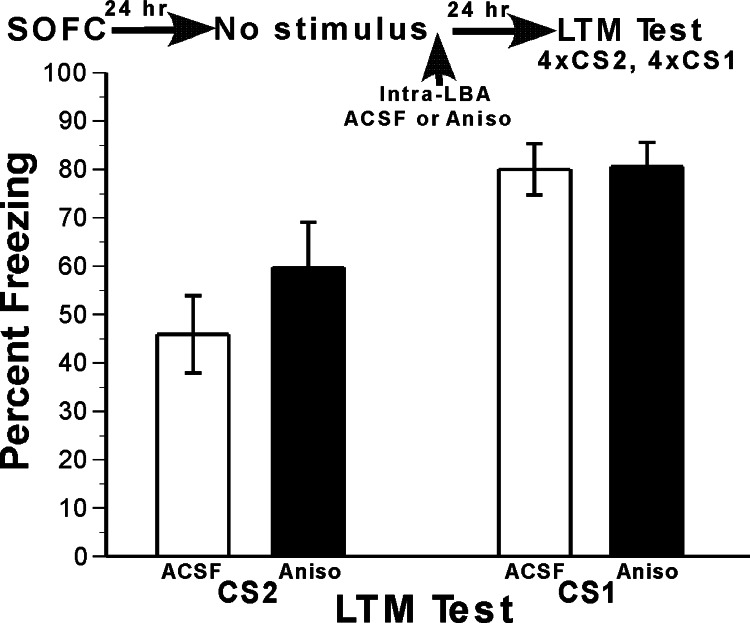
Memory consolidation refers to a process by which newly learned information is made resistant to disruption. Traditionally, consolidation has been viewed as an event that occurs once in the life of a memory. However, considerable evidence now indicates that consolidated memories, when reactivated through retrieval, become labile (susceptible to disruption) again and undergo reconsolidation. Because memories are often interrelated in complex associative networks rather than stored in isolation, a key question is whether reactivation of one memory makes associated memories labile in a way that requires reconsolidation. We tested this in rats by creating interlinked associative memories using a second-order fear-conditioning task. We found that directly reactivated memories become labile, but indirectly reactivated (i.e., associated) memories do not. This suggests that memory reactivation produces content-limited rather than wholesale changes in a memory and its associations and explains why each time a memory is retrieved and updated, the entire associative structure of the memory is not grossly altered.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- McGaugh J. L. Science. 2000;287:248–251. - PubMed
-
- Davis H. P., Squire L. R. Psychol. Bull. 1984;96:518–559. - PubMed
-
- Goelet P., Castellucci V. F., Schacher S., Kandel E. R. Nature. 1986;322:419–422. - PubMed
-
- Dudai Y. Annu. Rev. Psychol. 2004;55:51–86. - PubMed
-
- Sara S. J. Learn. Mem. 2000;7:73–84. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
