

Review
doi: 10.1513/pats.200510-109JH.
The aquaporin water channels
Affiliations
- PMID: 16493146
- PMCID: PMC2658677
- DOI: 10.1513/pats.200510-109JH
Item in Clipboard
Review
The aquaporin water channels
Proc Am Thorac Soc.
2006.
No abstract available
Figures

Discovery of aquaporin-1 (AQP1). Functional demonstration of water transport in X. laevis oocytes. Top panel: Control oocyte not expressing AQP1 (left) and test oocyte expressing AQP1 (right) 15 s after transfer to hypoosmotic culture medium. Middle panel: Same oocytes 3 min after transfer to hypoosmotic culture medium. The control oocyte failed to swell. The test oocyte swelled rapidly by osmosis and ruptured. Reprinted by permission from Preston GM, Carroll TP, Guggino WB, Agre P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 1992;256:385–387. Bottom panel: Jubilation expressed by Postdoctoral Fellow Gregory Preston 3 years after original report.

Discovery of aquaporin-1 (AQP1). Functional demonstration of water transport in X. laevis oocytes. Top panel: Control oocyte not expressing AQP1 (left) and test oocyte expressing AQP1 (right) 15 s after transfer to hypoosmotic culture medium. Middle panel: Same oocytes 3 min after transfer to hypoosmotic culture medium. The control oocyte failed to swell. The test oocyte swelled rapidly by osmosis and ruptured. Reprinted by permission from Preston GM, Carroll TP, Guggino WB, Agre P. Appearance of water channels in Xenopus oocytes expressing red cell CHIP28 protein. Science 1992;256:385–387. Bottom panel: Jubilation expressed by Postdoctoral Fellow Gregory Preston 3 years after original report.
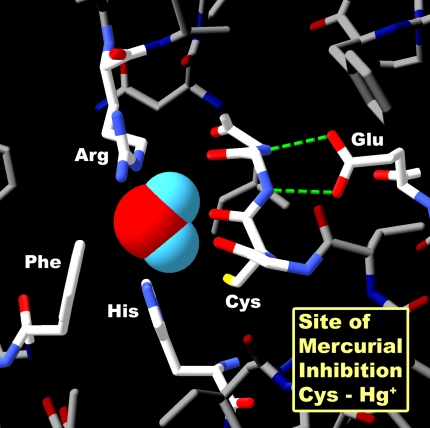
Structure of AQP1–Hg++ inhibitory site. Space-filling representation of a single water molecule in the narrowest point in the pore. Surrounding residues arginine-195 (Arg) and histidine-180 (His) provide positive charges that repel protons. Cysteine-189 (Cys) is the site of Hg++ inhibition. Phenylalanine-56 (Phe) and glutamic acid-142 (Glu) are other important residues in the pore region. Reprinted by permission from Kozono D, Yasui M, King LS, Agre P. Aquaporin water channels: atomic structure molecular dynamics meet clinical medicine. J Clin Invest 2002;109:1395–1399.
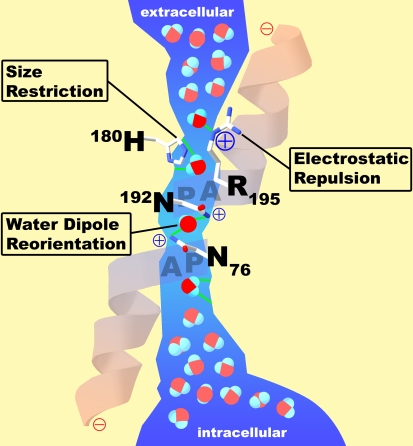
Functional schematic for water passage through AQP1. The extracellular vestibule and the intracellular vestibule of the channel contain water in bulk solution. They are connected by a 20-Å span where water molecules pass in single file. Barriers to the passage of protons are visible. Arginine-195 (R195) and histidine-180 (H180) provide fixed positive charges to repel proton passage. A single water molecule forms hydrogen bonds with the side chains of highly conserved asparagines-76 and -192 (N76 and N192). Partial positive charges are provided by the orientation of the two α helices that enter but do not entirely span the bilayer. Reprinted by permission from Kozono D, Yasui M, King LS, Agre P. Aquaporin water channels: atomic structure molecular dynamics meet clinical medicine. J Clin Invest 2002;109:1395–1399.
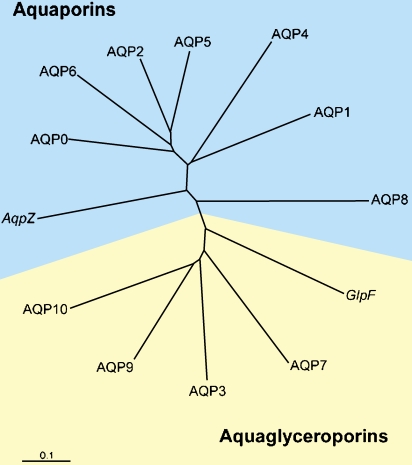
Human aquaporin repertoire. Two subsets of members exist: those permeated only by water (aquaporins) and those permeated by water plus glycerol (aquaglyceroporins). Included are the two members from Escherichia coli: AqpZ, a water channel, and GlpF, a glycerol transporter. Reprinted by permission from Kozono D, Yasui M, King LS, Agre P. Aquaporin water channels: atomic structure molecular dynamics meet clinical medicine. J Clin Invest 2002;109:1395–1399.
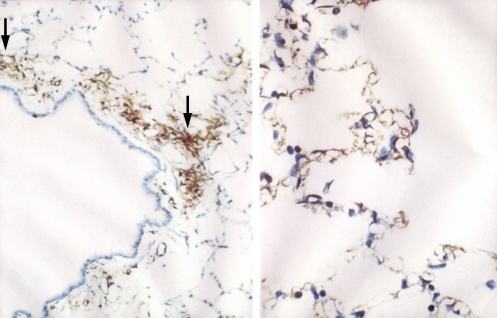
AQP1 in rat lung capillary endothelium. Left panel: Immunoperoxidase staining reveals AQP1 in capillary endothelium (arrows) in peribronchiolar distribution. Right panel: Immunoperoxidase staining reveals AQP1 in capillary endothelium in alveolar distribution. Reprinted by permission from Nielsen S, Smith BL, Christensen EI, Agre P. Distribution of the aquaporin CHIP in secretory and resorptive epithelia and capillary endothelia. Proc Natl Acad Sci 1993;90: 7275–7279.
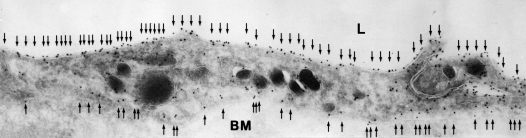
AQP1 in human lung capillary endothelium. Immunogold electron microscopic analysis reveals labeling of plasma membranes on lumenal surface (top) and ablumenal surface (bottom). Reprinted by permission from King LS, Nielsen S, Agre P, Brown RH. Decreased pulmonary vascular permeability in aquaporin-1-null humans. Proc Natl Acad Sci 2002;99: 1059–1063.
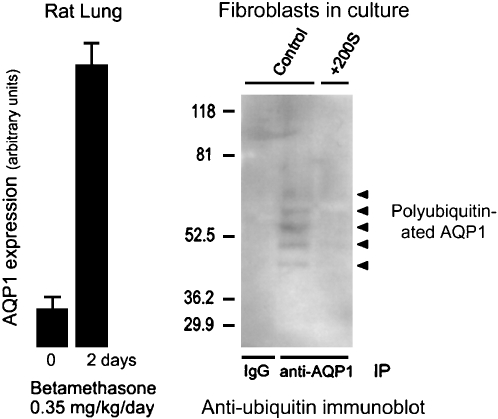
Dynamics of AQP1 expression. Left panel: Expression of AQP1 in rat lung is induced by corticosteroid exposure for 2 d. Reprinted by permission from King LS, Nielsen S, Agre P. Aquaporin-1 water channel protein in lung: ontogeny, steroid-induced expression, and distribution in rat. J Clin Invest 1996;97:2183–2191. Right panel: Level of AQP1 expression in cultured fibroblasts induced by hypertonic shock is regulated by degradation through the ubiquitin–proteosome pathway. Controls (left lane and center lane) denote fibroblasts grown in isotonic culture media producing extremely low levels of AQP1 accumulation. +200S denotes fibroblasts grown in culture medium made hypertonic by addition of 200 mOsm sorbitol. Anti-AQP1 immunoprecipitation (IP) was followed by antiubiquitin immunoblot. The left lane shows no signal in the control lacking immunoprecipitation control. The center lane shows polyubiquitination of AQP1 (ladder of reactive bands) in fibroblasts grown in isotonic culture media, thereby keeping AQP1 expression levels low. The right lane shows no polyubiquitination of AQP1 in fibroblasts grown in hypertonic culture medium, allowing AQP1 expression levels to rise to high levels. Reprinted by permission from Leitch V, Agre P, King LS. Altered ubiquitination and stability of aquaporin-1 in hypertonic stress. Proc Natl Acad Sci 2001;98:2894–2898.
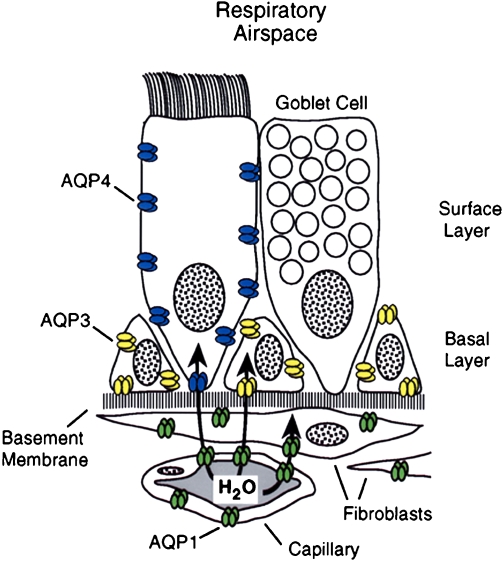
Aquaporins in respiratory epithelia. Upper panel: Schematic demonstrating absence of aquaporin expression in goblet cells, AQP4 in surface columnar cells, AQP3 in basal cells, and AQP1 in underlying fibroblasts and capillaries. Lower panels: Immunoperoxidase, brown staining, reveals aquaporins in rat tissue sections corresponding to nasopharyngeal surface epithelium. (A) AQP3, (B) AQP4, (C) AQP5 (no staining). and (D) AQP1. Immunoperoxidase, brown staining, reveals aquaporins in rat tissue sections corresponding to subepithelial glands. (E) AQP3 (no staining), (F) control (no staining,) and (G) AQP5 (staining of apical surface). Reprinted by permission from Nielsen S, King LS, Christensen BM, Agre P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am J Physiol Cell Physiol 1997; 273:C1549–C1561.
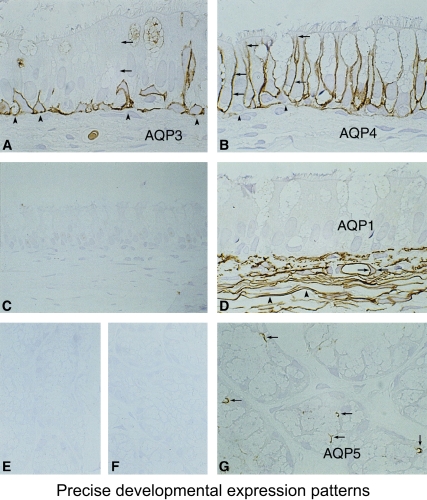
Aquaporins in respiratory epithelia. Upper panel: Schematic demonstrating absence of aquaporin expression in goblet cells, AQP4 in surface columnar cells, AQP3 in basal cells, and AQP1 in underlying fibroblasts and capillaries. Lower panels: Immunoperoxidase, brown staining, reveals aquaporins in rat tissue sections corresponding to nasopharyngeal surface epithelium. (A) AQP3, (B) AQP4, (C) AQP5 (no staining). and (D) AQP1. Immunoperoxidase, brown staining, reveals aquaporins in rat tissue sections corresponding to subepithelial glands. (E) AQP3 (no staining), (F) control (no staining,) and (G) AQP5 (staining of apical surface). Reprinted by permission from Nielsen S, King LS, Christensen BM, Agre P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am J Physiol Cell Physiol 1997; 273:C1549–C1561.
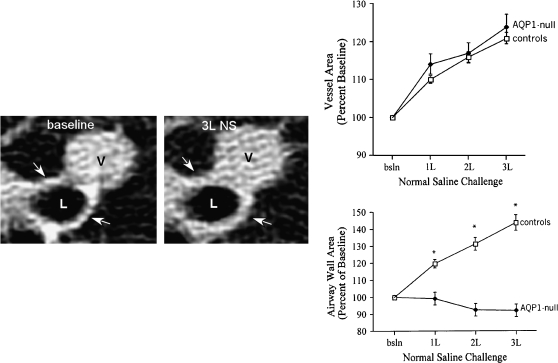
AQP1-null humans—pulmonary capillary defect. Left panels: High-resolution computed tomography scan of lung before (left) and after (right) infusion of 3 L of physiologic saline. The lumen of a 2-mm bronchiole is denoted (L) as well as an adjacent venule (V). After infusion of saline, the venule becomes engorged and the wall of the bronchiole becomes thickened due to accumulation of fluid (see arrows). Upper right panel: This was reproduced in multiple normal individuals. Lower right panel: When AQP1-null individuals were investigated, they did not accumulate fluid surrounding the bronchiole, since the vascular water permeability was reduced *p<0.001 between groups. Reprinted by permission from King LS, Nielsen S, Agre P, Brown RH. Decreased pulmonary vascular permeability in aquaporin-1-null humans. Proc Natl Acad Sci 2002;99:1059–1063.
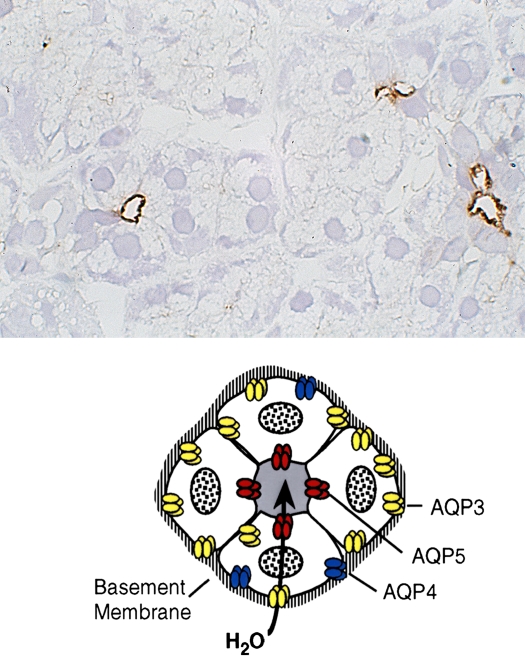
AQP5 in secretory glands. Drawings of secretory glands reveal AQP5, colored red, at the apical membrane (lower panel). The immunohistochemically stained tissue section of rat salivary gland reveals AQP5, stained brown, at the apical surface of three acini but not basolateral membranes (upper panel). Reprinted by permission from Nielsen S, King LS, Christensen BM, Agre P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am J Physiol Cell Physiol 1997;273:C1549–C1561.
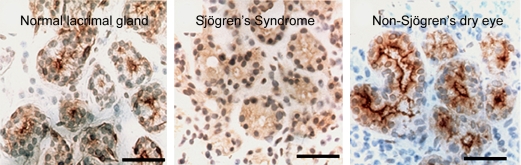
AQP5 in lacrimal glands—anti-AQP5 immunohistochemical staining of normal versus Sjögren's syndrome biopsies. Paraffin sections of human biopsies show AQP5, brown staining, in the apical membrane of normal (left panel) and non-Sjögren's dry eye (right panel). Center panel: The biopsy from a patient with Sjögren's syndrome reveals diffuse staining over the cell body indicating that a mistake in membrane trafficking had occurred. Studies by other investigators (Beroukas and colleagues) did not reveal abnormal AQP5 distribution but demonstrated AQP1 is reduced in myoepithelium (not shown). Reprinted by permission from Tsubota K, Hirai S, King LS, Agre P, Ishida N. Defective cellular trafficking of lacrimal gland aquaporin-5 is Sjögren's syndrome. Lancet 2001;357:688–689.
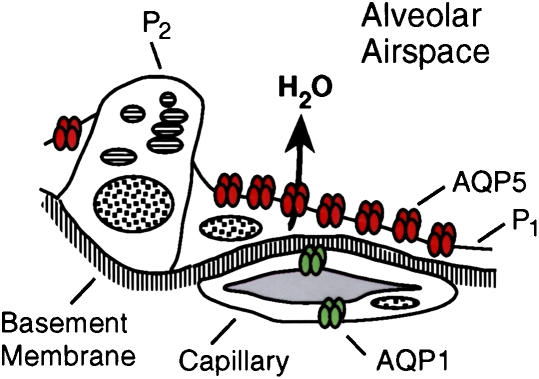
AQP5 in alveolar type 1 pneumocytes. Upper panel: Drawing of alveolar space reveals AQP5, colored red, in apical membrane of type 1 pneumocytes (P1) but not type 2 pneumocytes (P2). Middle and lower panels: Immunogold labeling of AQP5 at apical membranes denoted with arrows. ENDO = endothelium; EPI = epithelium. Reprinted by permission from Nielsen S, King LS, Christensen BM, Agre P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am J Physiol Cell Physiol 1997;273:C1549–C1561 (upper and middle panels); King LS, Nielsen S, Agre P. Respiratory aquaporins in lung inflammation: the night is young. Am J Respir Cell Mol Biol 2000;22:8–10 (lower panel).
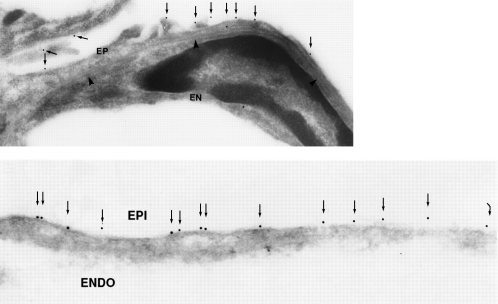
AQP5 in alveolar type 1 pneumocytes. Upper panel: Drawing of alveolar space reveals AQP5, colored red, in apical membrane of type 1 pneumocytes (P1) but not type 2 pneumocytes (P2). Middle and lower panels: Immunogold labeling of AQP5 at apical membranes denoted with arrows. ENDO = endothelium; EPI = epithelium. Reprinted by permission from Nielsen S, King LS, Christensen BM, Agre P. Aquaporins in complex tissues. II. Subcellular distribution in respiratory and glandular tissues of rat. Am J Physiol Cell Physiol 1997;273:C1549–C1561 (upper and middle panels); King LS, Nielsen S, Agre P. Respiratory aquaporins in lung inflammation: the night is young. Am J Respir Cell Mol Biol 2000;22:8–10 (lower panel).
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases