

Interleukin-5 (IL-5) augments the progression of liver fibrosis by regulating IL-13 activity
- PMID: 16495517
- PMCID: PMC1418671
- DOI: 10.1128/IAI.74.3.1471-1479.2006
Interleukin-5 (IL-5) augments the progression of liver fibrosis by regulating IL-13 activity
Abstract
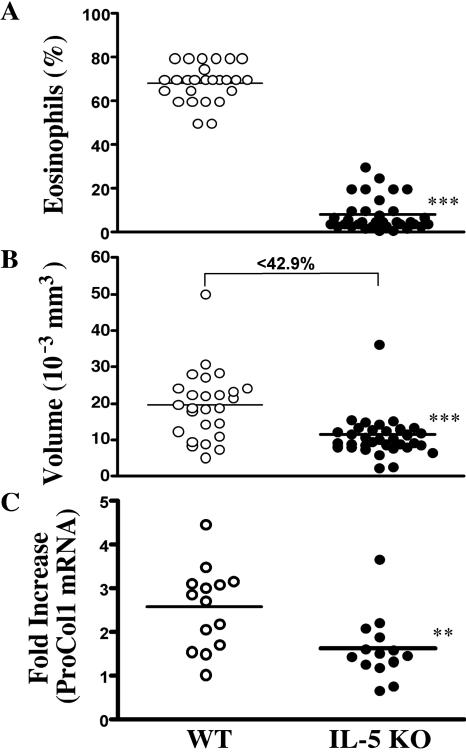
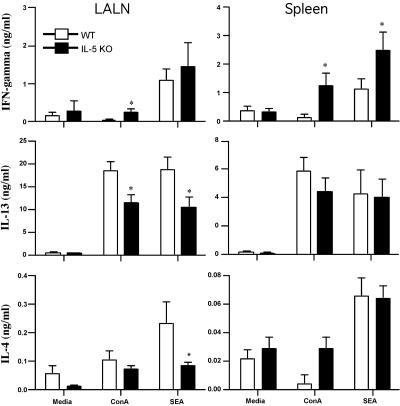
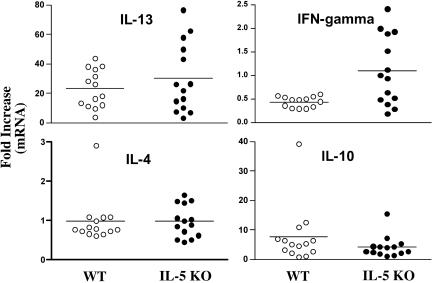
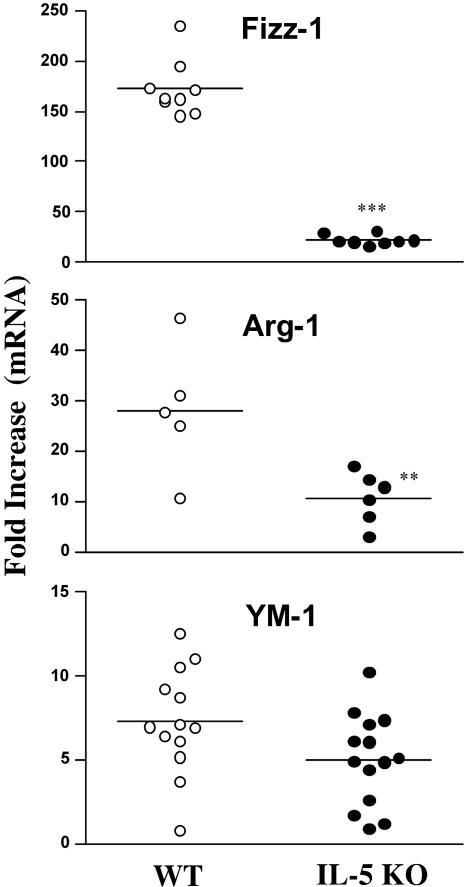
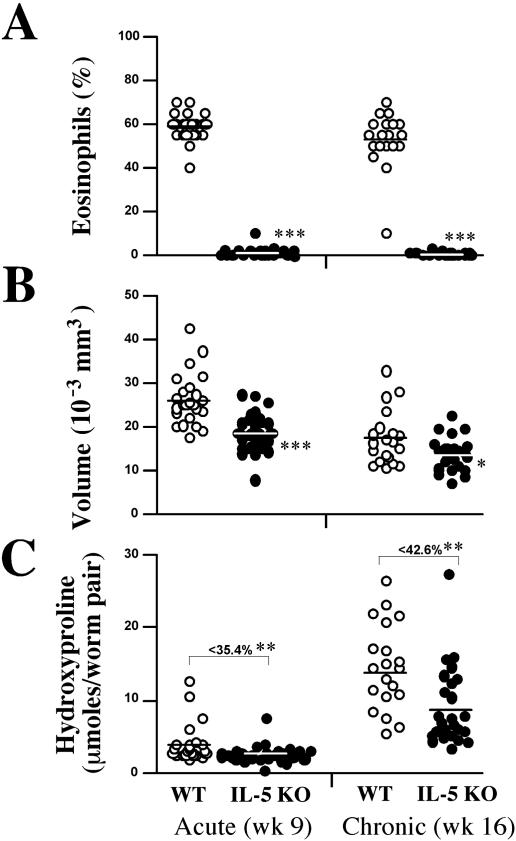
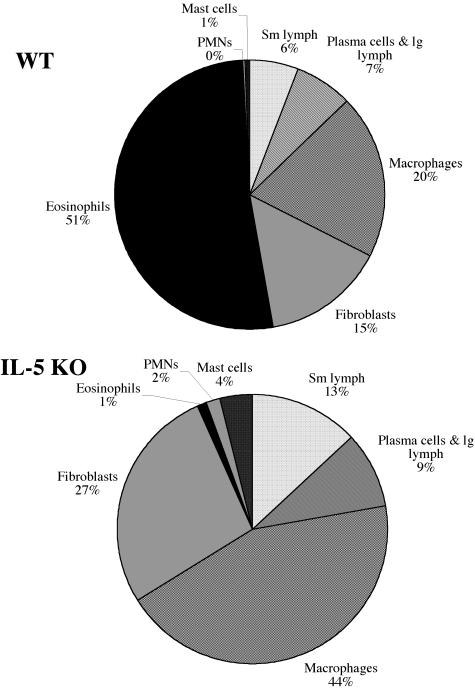
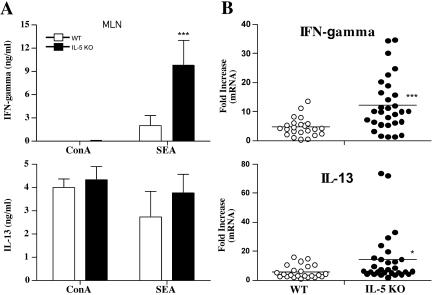
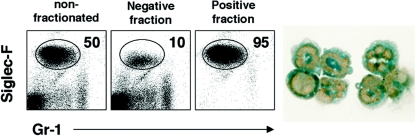
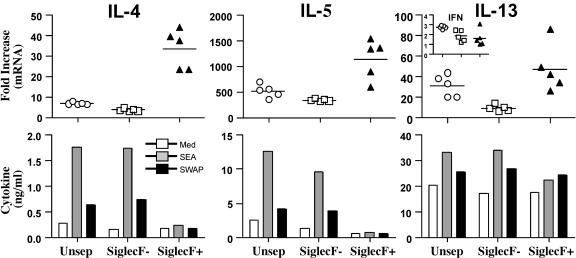
Eosinophils are frequently found in increased numbers in a variety of chronic fibrotic diseases; however, their role in the development of hepatic fibrosis has not been dissected in vivo. Here, we used interleukin-5 (IL-5) knockout (KO) mice to determine whether eosinophils contribute to the progressive liver fibrosis that develops in response to chronic Schistosoma mansoni infection. Although infection intensities were similar in C57BL/6 and IL-5 KO mice, the average size of granulomas was significantly smaller in both acutely and chronically infected IL-5 KO mice. Their granulomas were also completely devoid of eosinophils. In addition, the knockout mice displayed over a 40% reduction in hepatic fibrosis by week 16 postinfection. The reduced fibrosis was associated with increased production of the antifibrotic cytokine gamma interferon. Moreover, although IL-13 production did not decrease consistently in the absence of IL-5, IL-13-triggered responses were substantially reduced in the granulomatous tissues. This was confirmed by analyzing the expression of several genes associated with alternative macrophage activation, including arginase 1, Fizz-1, and YM-1. Importantly, all of these IL-13-regulated genes have been linked with the mechanisms of wound healing and fibrosis. In addition to IL-5 polarizing the antigen-specific CD4+ Th2 cell response, we found that granuloma eosinophils were themselves a significant source of IL-13. Thus, by producing profibrotic mediators and polarizing the Th2 response, these findings illustrate both direct and indirect roles for eosinophils and IL-5 in the pathogenesis of schistosomiasis-induced liver fibrosis. Thus, inhibiting the activity of IL-5 or eosinophils may prove effective for a variety of chronic fibrotic diseases.
Figures
Similar articles
-
Lack of antigen-specific Th1 response alters granuloma formation and composition in Schistosoma mansoni-infected MyD88-/- mice.Eur J Immunol. 2005 Nov;35(11):3248-57. doi: 10.1002/eji.200526273. Eur J Immunol. 2005. PMID: 16276483
-
The granulomatous response in murine Schistosomiasis mansoni does not switch to Th1 in IL-4-deficient C57BL/6 mice.J Immunol. 1996 Nov 15;157(10):4546-53. J Immunol. 1996. PMID: 8906833
-
Macrophage migration inhibitory factor is critical to interleukin-5-driven eosinophilopoiesis and tissue eosinophilia triggered by Schistosoma mansoni infection.FASEB J. 2009 Apr;23(4):1262-71. doi: 10.1096/fj.08-124248. Epub 2008 Dec 16. FASEB J. 2009. PMID: 19088181
-
The role of cytokines in the formation of the schistosome egg granuloma.Immunobiology. 1994 Oct;191(4-5):441-50. doi: 10.1016/S0171-2985(11)80450-X. Immunobiology. 1994. PMID: 7713558 Review.
-
Th1 and Th2 responses regulate experimental lung granuloma development.Sarcoidosis Vasc Diffuse Lung Dis. 1996 Sep;13(2):120-8. Sarcoidosis Vasc Diffuse Lung Dis. 1996. PMID: 8893380 Review.
Cited by
-
Cytokine Pattern of T Lymphocytes in Acute Schistosomiasis mansoni Patients following Treated Praziquantel Therapy.J Parasitol Res. 2013;2013:909134. doi: 10.1155/2013/909134. Epub 2013 Jan 20. J Parasitol Res. 2013. PMID: 23401741 Free PMC article.
-
Hepatic macrophages in liver fibrosis: pathogenesis and potential therapeutic targets.BMJ Open Gastroenterol. 2016 May 25;3(1):e000079. doi: 10.1136/bmjgast-2016-000079. eCollection 2016. BMJ Open Gastroenterol. 2016. PMID: 27252881 Free PMC article.
-
Immunopathology of schistosomiasis.Immunol Cell Biol. 2007 Feb-Mar;85(2):148-54. doi: 10.1038/sj.icb.7100014. Epub 2006 Dec 12. Immunol Cell Biol. 2007. PMID: 17160074 Free PMC article. Review.
-
Macrophages: master regulators of inflammation and fibrosis.Semin Liver Dis. 2010 Aug;30(3):245-57. doi: 10.1055/s-0030-1255354. Epub 2010 Jul 21. Semin Liver Dis. 2010. PMID: 20665377 Free PMC article. Review.
-
A major ecological niche of eosinophils in evolving Schistosoma granulomas challenges the eosinophil view as "helminth killer" cells.Sci Adv. 2025 Jun 13;11(24):eadt2779. doi: 10.1126/sciadv.adt2779. Epub 2025 Jun 11. Sci Adv. 2025. PMID: 40498832 Free PMC article.
References
-
- Brunet, L. R., E. A. Sabin, A. W. Cheever, M. A. Kopf, and E. J. Pearce. 1999. Interleukin 5 (IL-5) is not required for expression of a Th2 response or host resistance mechanisms during murine schistosomiasis mansoni but does play a role in development of IL-4-producing non-T, non-B cells. Infect. Immun. 67:3014-3018. - PMC - PubMed
-
- Bystrom, J., T. A. Wynn, J. B. Domachowske, and H. F. Rosenberg. 2004. Gene microarray analysis reveals interleukin-5-dependent transcriptional targets in mouse bone marrow. Blood 103:868-877. [Online.] - PubMed
-
- Capron, M. 1992. Dual function of eosinophils in pathogenesis and protective immunity against parasites. Mem. Inst. Oswaldo Cruz 87:83-89. - PubMed
-
- Cheever, A. W., M. E. Williams, T. A. Wynn, F. D. Finkelman, R. A. Seder, T. M. Cox, S. Hieny, P. Caspar, and A. Sher. 1994. Anti-IL-4 treatment of Schistosoma mansoni-infected mice inhibits development of T cells and non-B, non-T cells expressing Th2 cytokines while decreasing egg-induced hepatic fibrosis. J. Immunol. 153:753-759. - PubMed
-
- Cheever, A. W., Y. Xu, J. G. Macedonia, T. Cox, S. Hieny, and A. Sher. 1992. The role of cytokines in the pathogenesis of hepatic granulomatous disease in Schistosoma mansoni infected mice. Mem. Inst. Oswaldo Cruz 87:81-85. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
