

Identification of fibronectin-binding proteins in Mycoplasma gallisepticum strain R
- PMID: 16495551
- PMCID: PMC1418676
- DOI: 10.1128/IAI.74.3.1777-1785.2006
Identification of fibronectin-binding proteins in Mycoplasma gallisepticum strain R
Abstract
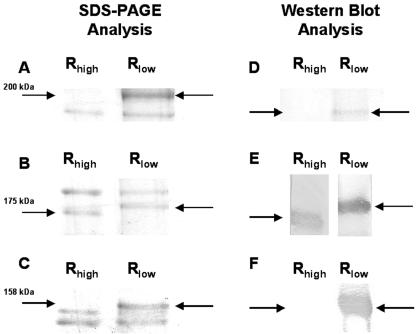
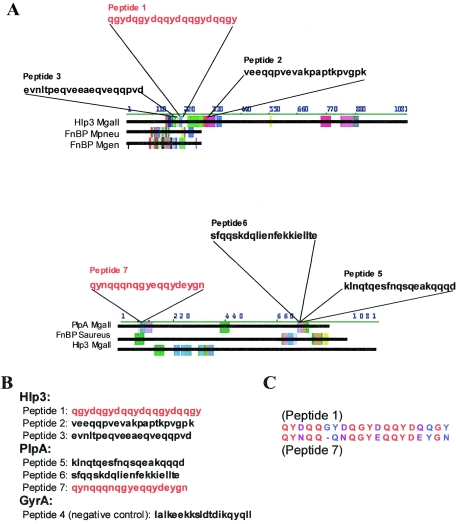
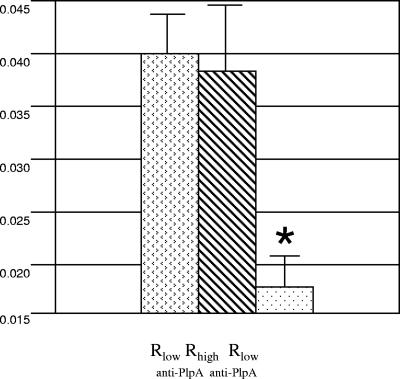
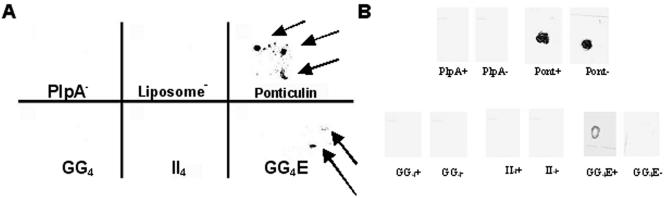
We have determined that virulent Mycoplasma gallisepticum strain Rlow is capable of binding the extracellular matrix protein fibronectin. Fibronectin was found to be present in M. gallisepticum Rlow protein extracts by Western blotting and peptide sequencing. Mycoplasma gallisepticum Rhigh, the attenuated, high-passage derivative of Rlow, is deficient in this ability. MGA_1199, the M. gallisepticum homologue of the cytadherence-associated protein P65 from Mycoplasma pneumoniae, and MGA_0928, the M. gallisepticum homologue of the M. pneumoniae cytoskeletal protein HMW3, were identified as fibronectin-binding proteins. Peptides from the regions of MGA_1199 and MGA_0928 exhibiting the highest degree of homology with known fibronectin-binding proteins were shown to bind the gelatin/heparin-binding domain of fibronectin. MGA_1199 and MGA_0928 were shown to be absent and aberrant, respectively, in Rhigh, explaining its lack of fibronectin-binding capability. Consistent with its M. pneumoniae counterpart, MGA_1199 (renamed PlpA) was demonstrated to be surface exposed, despite a lack of classical membrane-spanning domains. Due to its demonstrated topology and the strength of interaction between its binding peptide and fibronectin, we propose that PlpA functions as a fibronectin-binding protein in vivo and may possess atypical transmembrane domains.
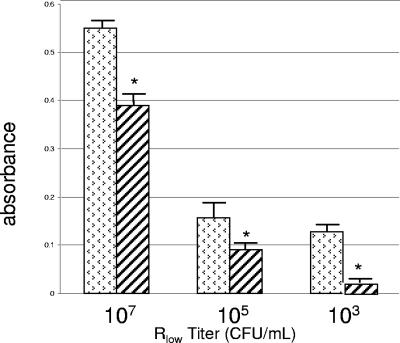
Figures


Similar articles
-
Mycoplasma gallisepticum in vivo induced antigens expressed during infection in chickens.Vet Microbiol. 2015 Feb 25;175(2-4):265-74. doi: 10.1016/j.vetmic.2014.12.007. Epub 2014 Dec 18. Vet Microbiol. 2015. PMID: 25575879
-
The Mycoplasma gallisepticum OsmC-like protein MG1142 resides on the cell surface and binds heparin.Microbiology (Reading). 2007 May;153(Pt 5):1455-1463. doi: 10.1099/mic.0.2006/004937-0. Microbiology (Reading). 2007. PMID: 17464059
-
Mycoplasma gallisepticum hemagglutinin V1hA, pyruvate dehydrogenase PdhA, lactate dehydrogenase, and elongation factor Tu share epitopes with Mycoplasma imitans homologues.Avian Dis. 2005 Dec;49(4):507-13. doi: 10.1637/7377-051105R.1. Avian Dis. 2005. PMID: 16404991
-
Metabolite profiling of Mycoplasma gallisepticum mutants, combined with bioinformatic analysis, can reveal the likely functions of virulence-associated genes.Vet Microbiol. 2018 Sep;223:160-167. doi: 10.1016/j.vetmic.2018.08.001. Epub 2018 Aug 2. Vet Microbiol. 2018. PMID: 30173742
-
The future of whole-cell modeling.Curr Opin Biotechnol. 2014 Aug;28:111-5. doi: 10.1016/j.copbio.2014.01.012. Epub 2014 Feb 17. Curr Opin Biotechnol. 2014. PMID: 24556244 Free PMC article. Review.
Cited by
-
An emerging role for cyclic dinucleotide phosphodiesterase and nanoRNase activities in Mycoplasma bovis: Securing survival in cell culture.PLoS Pathog. 2020 Jun 29;16(6):e1008661. doi: 10.1371/journal.ppat.1008661. eCollection 2020 Jun. PLoS Pathog. 2020. PMID: 32598377 Free PMC article.
-
Infection strategies of mycoplasmas: Unraveling the panoply of virulence factors.Virulence. 2021 Dec;12(1):788-817. doi: 10.1080/21505594.2021.1889813. Virulence. 2021. PMID: 33704021 Free PMC article. Review.
-
Variable Lipoprotein Hemagglutinin A Gene (vlhA) Expression in Variant Mycoplasma gallisepticum Strains In Vivo.Infect Immun. 2018 Oct 25;86(11):e00524-18. doi: 10.1128/IAI.00524-18. Print 2018 Nov. Infect Immun. 2018. PMID: 30181349 Free PMC article.
-
Detection and differentiation of avian mycoplasmas by surface-enhanced Raman spectroscopy based on a silver nanorod array.Appl Environ Microbiol. 2012 Mar;78(6):1930-5. doi: 10.1128/AEM.07419-11. Epub 2011 Dec 30. Appl Environ Microbiol. 2012. PMID: 22210215 Free PMC article.
-
A Computational Reverse Vaccinology Approach for the Design and Development of Multi-Epitopic Vaccine Against Avian Pathogen Mycoplasma gallisepticum.Front Vet Sci. 2021 Oct 26;8:721061. doi: 10.3389/fvets.2021.721061. eCollection 2021. Front Vet Sci. 2021. PMID: 34765664 Free PMC article.
References
-
- Balish, M. F., and D. C. Krause. 2002. Cytadherence and the cytoskeleton, p. 491-518. In S. Razin and R. Herrmann (ed.), Molecular biology and pathogenicity of mycoplasmas. Kluwer Academic/Plenum Publishers, New York, N.Y.
-
- Baseggio, N., M. D. Glew, P. F. Markham, K. G. Whithear, and G. F. Browning. 1996. Size and genomic location of the pMGA multigene family of Mycoplasma gallisepticum. Microbiology 142:1429-1435. - PubMed
-
- Bordier, C. 1981. Phase separation of integral membrane proteins in Triton X-114 solution. J. Biol. Chem. 256:1604-1607. - PubMed
-
- Cheng, H. C., M. Abdel-Ghany, R. C. Elble, and B. U. Pauli. 1998. Lung endothelial dipeptidyl peptidase IV promotes adhesion and metastasis of rat breast cancer cells via tumor cell surface-associated fibronectin. J. Biol. Chem. 273:24207-24215. - PubMed
-
- Chin, R. P., B. M. Daft, C. U. Meteyer, and R. Yamamoto. 1991. Meningoencephalitis in commercial meat turkeys associated with Mycoplasma gallisepticum. Avian Dis. 35:986-993. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
