

Role for the CD28 molecule in the control of Coxiella burnetii infection
- PMID: 16495554
- PMCID: PMC1418619
- DOI: 10.1128/IAI.74.3.1800-1808.2006
Role for the CD28 molecule in the control of Coxiella burnetii infection
Abstract
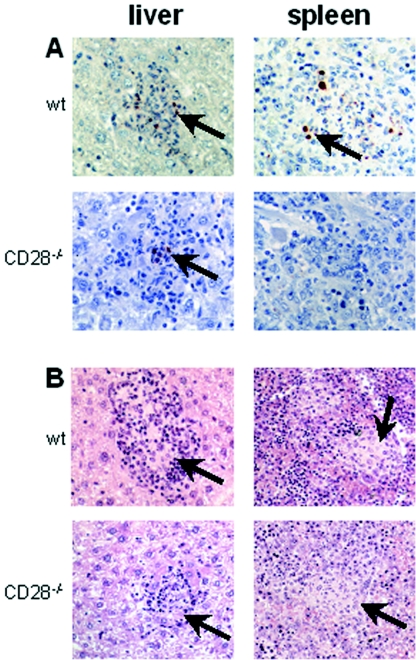
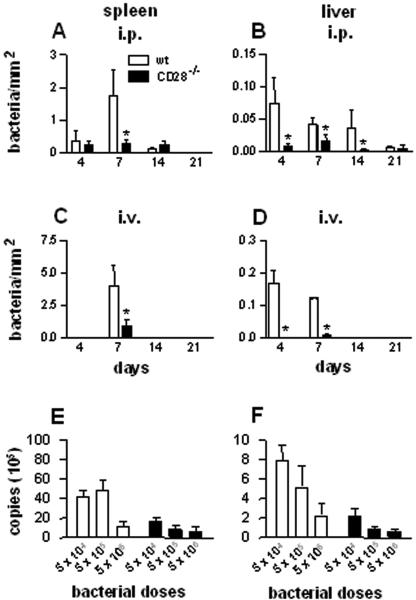
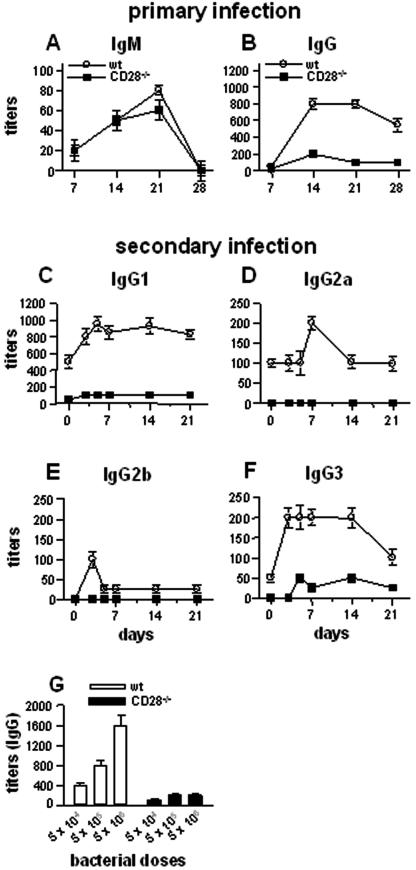
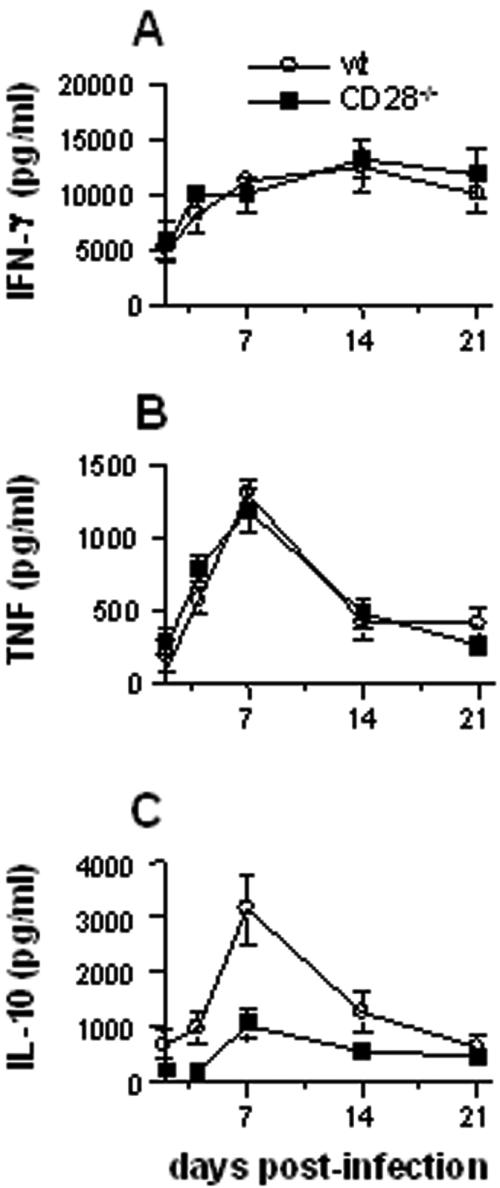
Q fever is an infectious disease caused by Coxiella burnetii, an obligate intracellular bacterium that replicates in macrophages. As cell-mediated immune response to microbial pathogens requires signals mediated by T-cell receptors and costimulatory molecules such as CD28, we wondered if CD28 is involved in protection against C. burnetii infection. CD28-deficient (CD28-/-) mice were inoculated with C. burnetii by intraperitoneal and intravenous routes. With both wild-type and CD28-/- mice, C. burnetii organisms were detected exclusively in spleen and liver. The antibody response against C. burnetii was impaired in CD28-/- animals, but, surprisingly, the lack of CD28 decreased C. burnetii burden in the infected tissues, whatever the manner of inoculation of bacteria. The CD28 deficiency had no effect on either granuloma formation, which reflects cell-mediated immunity against C. burnetii, or the production of gamma interferon and tumor necrosis factor, two cytokines known to be involved in granuloma formation. On the other hand, the production of interleukin-10 (IL-10) by peritoneal macrophages was highly impaired in CD28-/- mice. The results suggest that CD28 initiates a signal that favors C. burnetii replication through the modulation of the IL-10 pathway.
Figures
Similar articles
-
Vanin-1 controls granuloma formation and macrophage polarization in Coxiella burnetii infection.Eur J Immunol. 2007 Jan;37(1):24-32. doi: 10.1002/eji.200636054. Eur J Immunol. 2007. PMID: 17163446
-
IFN-gamma-mediated control of Coxiella burnetii survival in monocytes: the role of cell apoptosis and TNF.J Immunol. 1999 Feb 15;162(4):2259-65. J Immunol. 1999. PMID: 9973502
-
TLR2 is necessary to inflammatory response in Coxiella burnetii infection.Ann N Y Acad Sci. 2005 Dec;1063:161-6. doi: 10.1196/annals.1355.025. Ann N Y Acad Sci. 2005. PMID: 16481508
-
Intracellular life of Coxiella burnetii in macrophages.Ann N Y Acad Sci. 2009 May;1166:55-66. doi: 10.1111/j.1749-6632.2009.04515.x. Ann N Y Acad Sci. 2009. PMID: 19538264 Review.
-
Coxiella burnetii infection.Ann N Y Acad Sci. 2005 Dec;1063:105-14. doi: 10.1196/annals.1355.018. Ann N Y Acad Sci. 2005. PMID: 16481501 Review.
Cited by
-
A fatal case of disseminated chronic Q fever: a case report and brief review of the literature.Infection. 2016 Oct;44(5):677-82. doi: 10.1007/s15010-016-0884-0. Epub 2016 Mar 3. Infection. 2016. PMID: 26940462 Free PMC article. Review.
-
CD28 homodimer interface mimetic peptide acts as a preventive and therapeutic agent in models of severe bacterial sepsis and gram-negative bacterial peritonitis.J Infect Dis. 2015 Mar 15;211(6):995-1003. doi: 10.1093/infdis/jiu556. Epub 2014 Oct 9. J Infect Dis. 2015. PMID: 25305323 Free PMC article.
-
An experimental mouse model to establish Tropheryma whipplei as a diarrheal agent.J Infect Dis. 2011 Jul 1;204(1):44-50. doi: 10.1093/infdis/jir219. J Infect Dis. 2011. PMID: 21628657 Free PMC article.
-
B7-H4 Treatment of T Cells Inhibits ERK, JNK, p38, and AKT Activation.PLoS One. 2012;7(1):e28232. doi: 10.1371/journal.pone.0028232. Epub 2012 Jan 4. PLoS One. 2012. PMID: 22238573 Free PMC article.
-
Early cytokine and antibody responses against Coxiella burnetii in aerosol infection of BALB/c mice.Diagn Microbiol Infect Dis. 2015 Apr;81(4):234-9. doi: 10.1016/j.diagmicrobio.2014.12.008. Epub 2014 Dec 30. Diagn Microbiol Infect Dis. 2015. PMID: 25618420 Free PMC article.
References
-
- Andreasen, S. O., J. E. Christensen, O. Marker, and A. R. Thomsen. 2000. Role of CD40 ligand and CD28 in induction and maintenance of antiviral CD8+ effector T cell responses. J. Immunol. 164:3689-3697. - PubMed
-
- Andres, P. G., K. C. Howland, A. Nirula, L. P. Kane, L. Barron, D. Dresnek, A. Sadra, J. Imboden, A. Weiss, and A. K. Abbas. 2004. Distinct regions in the CD28 cytoplasmic domain are required for T helper type 2 differentiation. Nat. Immunol. 5:435-442. - PubMed
-
- Barton, G. M., and R. Medzhitov. 2002. Control of adaptive immune responses by Toll-like receptors. Curr. Opin. Immunol. 14:380-383. - PubMed
-
- Beck, J. M., M. B. Blackmon, C. M. Rose, S. L. Kimzey, A. M. Preston, and J. M. Green. 2003. T cell costimulatory molecule function determines susceptibility to infection with Pneumocystis carinii in mice. J. Immunol. 171:1969-1977. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
