

AtCyp59 is a multidomain cyclophilin from Arabidopsis thaliana that interacts with SR proteins and the C-terminal domain of the RNA polymerase II
- PMID: 16497658
- PMCID: PMC1421086
- DOI: 10.1261/rna.2226106
AtCyp59 is a multidomain cyclophilin from Arabidopsis thaliana that interacts with SR proteins and the C-terminal domain of the RNA polymerase II
Abstract
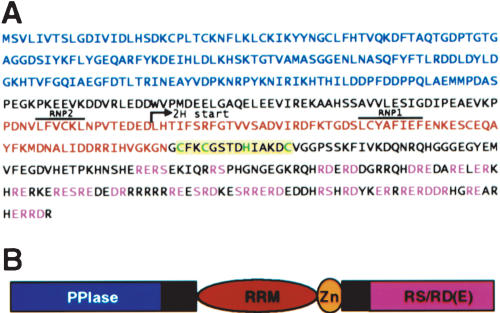
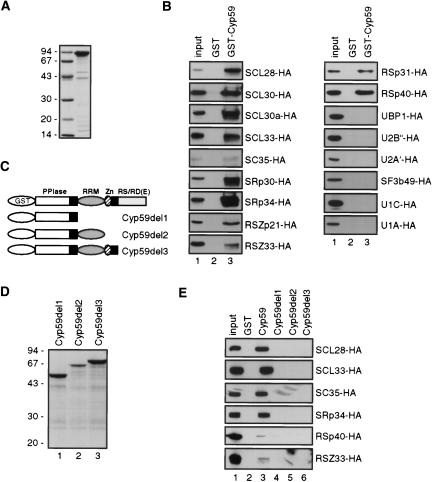
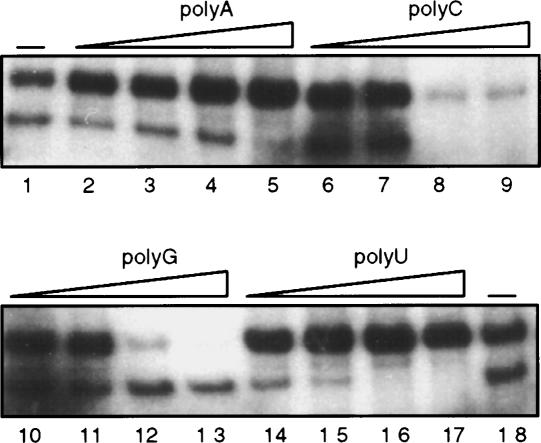
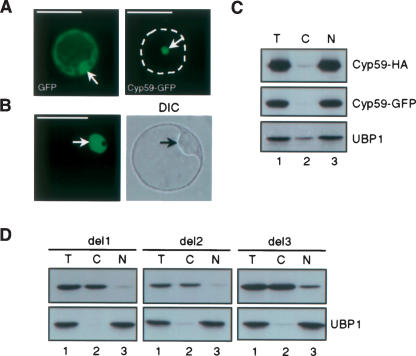
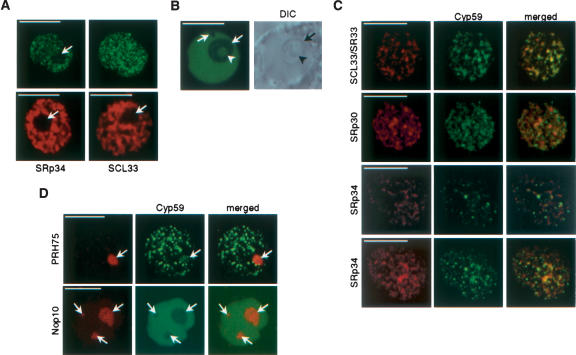
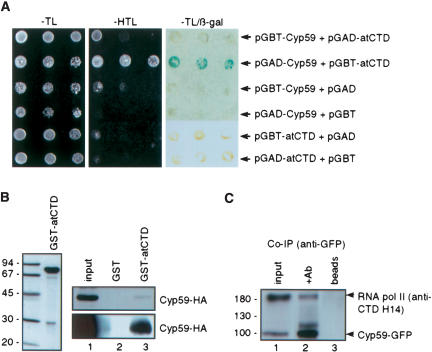
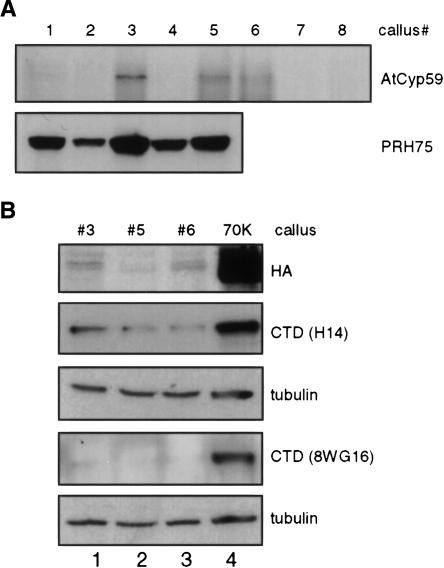
AtCyp59 and its orthologs from different organisms belong to a family of modular proteins consisting of a peptidyl-prolyl cis-trans isomerase (PPIase) domain, followed by an RNA recognition motif (RRM), and a C-terminal domain enriched in charged amino acids. AtCyp59 was identified in a yeast two-hybrid screen as an interacting partner of the Arabidopsis SR protein SCL33/SR33. The interaction with SCL33/SR33 and with a majority of Arabidopsis SR proteins was confirmed by in vitro pull-down assays. Consistent with these interactions, AtCyp59 localizes to the cell nucleus, but it does not significantly colocalize with SR proteins in nuclear speckles. Rather, it shows a punctuate localization pattern resembling transcription sites. Indeed, by using yeast two-hybrid, in vitro pull-down, and immunoprecipitation assays, we found that AtCyp59 interacts with the C-terminal domain (CTD) of the largest subunit of RNA polymerase II. Ectopic expression of the tagged protein in Arabidopsis cell suspension resulted in highly reduced growth that is most probably due to reduced phosphorylation of the CTD. Together our data suggest a possible function of AtCyp59 in activities connecting transcription and pre-mRNA processing. We discuss our data in the context of a dynamic interplay between transcription and pre-mRNA processing.
Figures

Similar articles
-
Identification of RNA targets for the nuclear multidomain cyclophilin atCyp59 and their effect on PPIase activity.Nucleic Acids Res. 2013 Feb 1;41(3):1783-96. doi: 10.1093/nar/gks1252. Epub 2012 Dec 16. Nucleic Acids Res. 2013. PMID: 23248006 Free PMC article.
-
Rct1, a nuclear RNA recognition motif-containing cyclophilin, regulates phosphorylation of the RNA polymerase II C-terminal domain.Mol Cell Biol. 2007 May;27(10):3601-11. doi: 10.1128/MCB.02187-06. Epub 2007 Mar 5. Mol Cell Biol. 2007. PMID: 17339332 Free PMC article.
-
Arabidopsis thaliana PRP40s are RNA polymerase II C-terminal domain-associating proteins.Arch Biochem Biophys. 2009 Apr 1;484(1):30-8. doi: 10.1016/j.abb.2009.01.004. Epub 2009 Jan 14. Arch Biochem Biophys. 2009. PMID: 19467629
-
The cyclophilins.Genome Biol. 2005;6(7):226. doi: 10.1186/gb-2005-6-7-226. Epub 2005 Jun 27. Genome Biol. 2005. PMID: 15998457 Free PMC article. Review.
-
Versatility of Cyclophilins in Plant Growth and Survival: A Case Study in Arabidopsis.Biomolecules. 2019 Jan 10;9(1):20. doi: 10.3390/biom9010020. Biomolecules. 2019. PMID: 30634678 Free PMC article. Review.
Cited by
-
Cyclophilins: proteins in search of function.Plant Signal Behav. 2013 Jan;8(1):e22734. doi: 10.4161/psb.22734. Epub 2012 Nov 3. Plant Signal Behav. 2013. PMID: 23123451 Free PMC article. Review.
-
Multiple abiotic stress responsive rice cyclophilin: (OsCYP-25) mediates a wide range of cellular responses.Commun Integr Biol. 2013 Sep 1;6(5):e25260. doi: 10.4161/cib.25260. Epub 2013 Jun 21. Commun Integr Biol. 2013. PMID: 24265852 Free PMC article.
-
The Conserved RNA Binding Cyclophilin, Rct1, Regulates Small RNA Biogenesis and Splicing Independent of Heterochromatin Assembly.Cell Rep. 2017 Jun 20;19(12):2477-2489. doi: 10.1016/j.celrep.2017.05.086. Cell Rep. 2017. PMID: 28636937 Free PMC article.
-
Regulatory Network of Serine/Arginine-Rich (SR) Proteins: The Molecular Mechanism and Physiological Function in Plants.Int J Mol Sci. 2022 Sep 5;23(17):10147. doi: 10.3390/ijms231710147. Int J Mol Sci. 2022. PMID: 36077545 Free PMC article. Review.
-
Genome-Wide Identification and Characterization of the Cyclophilin Gene Family in the Nematophagous Fungus Purpureocillium lilacinum.Int J Mol Sci. 2019 Jun 18;20(12):2978. doi: 10.3390/ijms20122978. Int J Mol Sci. 2019. PMID: 31216716 Free PMC article.
References
-
- Ali, G.S., Golovkin, M., and Reddy, A.S. 2003. Nuclear localization and in vivo dynamics of a plant-specific serine/arginine-rich protein. Plant J. 36: 883–893. - PubMed
-
- Bentley, D. 2002. The mRNA assembly line: Transcription and processing machines in the same factory. Curr. Opin. Cell Biol. 14: 336–342. - PubMed
-
- Burge, C.B., Tuschl, T., and Sharp, P.A. 1999. Splicing of precursors to mRNAs by the spliceosome. In The RNA world (eds. R.F. Gesteland et al.), pp. 525–560. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials