

Kinetics of force recovery following length changes in active skinned single fibres from rabbit psoas muscle: analysis and modelling of the late recovery phase
- PMID: 16497718
- PMCID: PMC1779726
- DOI: 10.1113/jphysiol.2005.103382
Kinetics of force recovery following length changes in active skinned single fibres from rabbit psoas muscle: analysis and modelling of the late recovery phase
Abstract
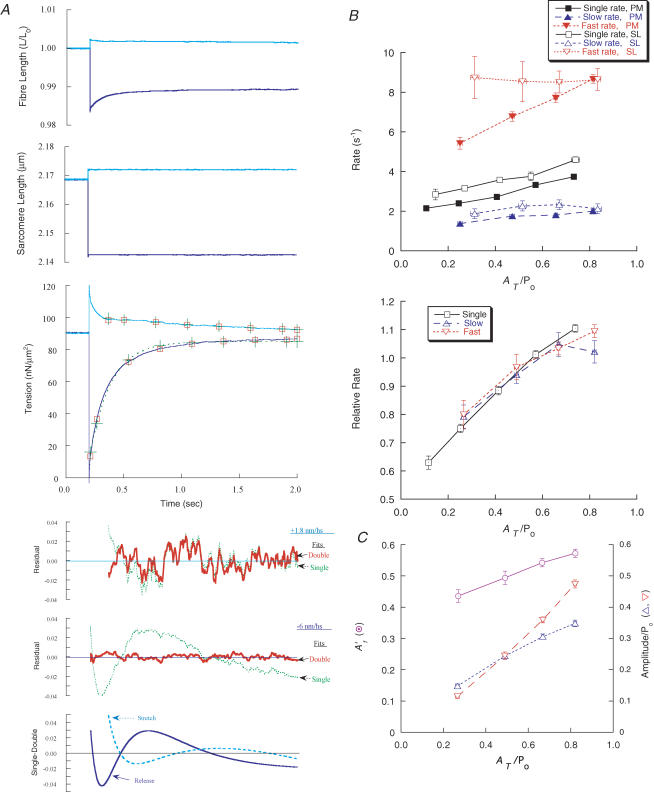
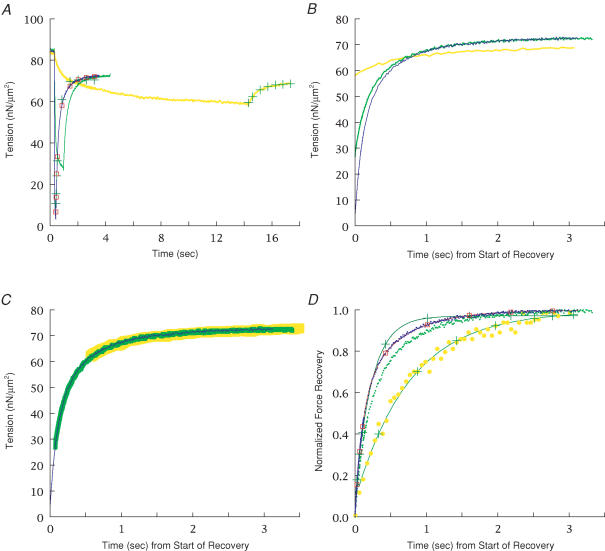
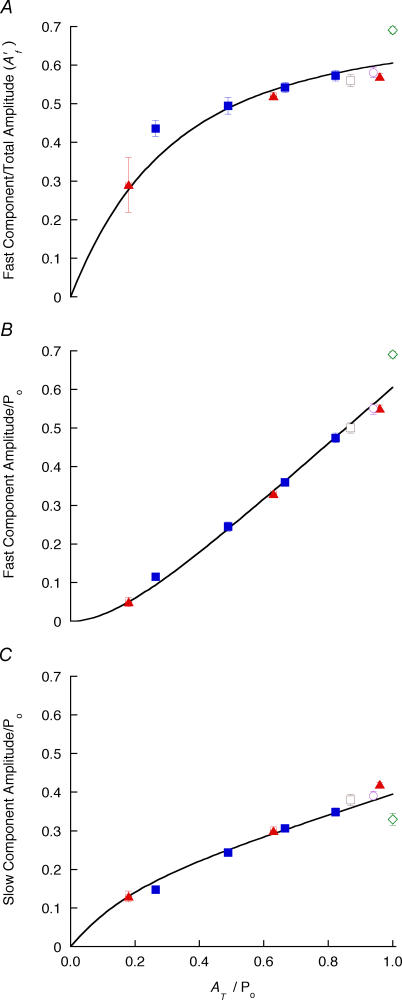
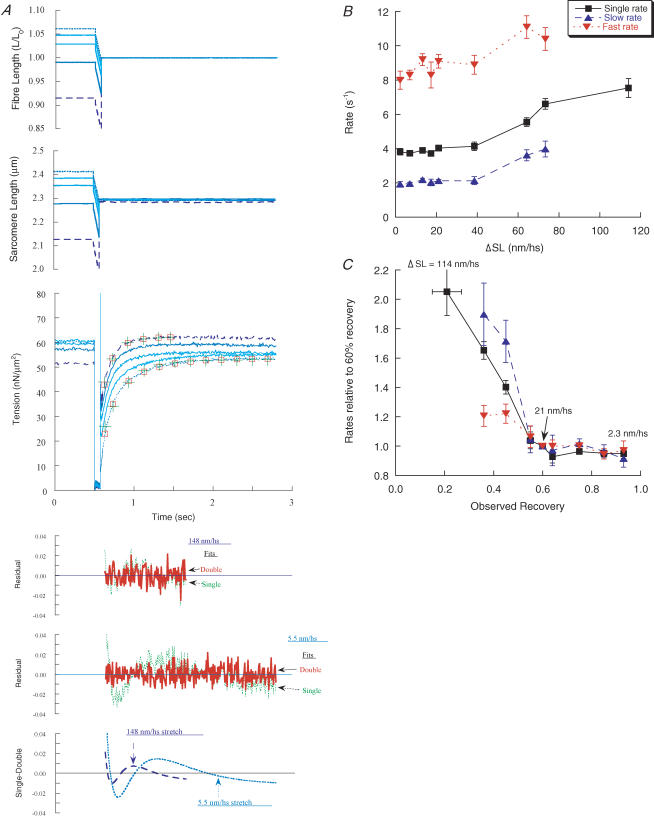
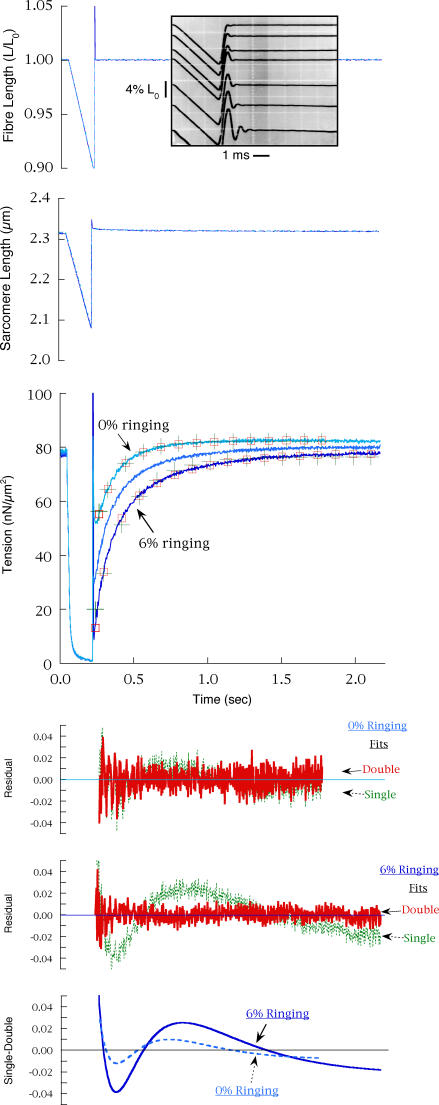
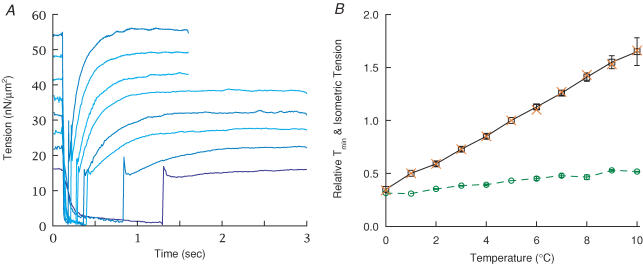
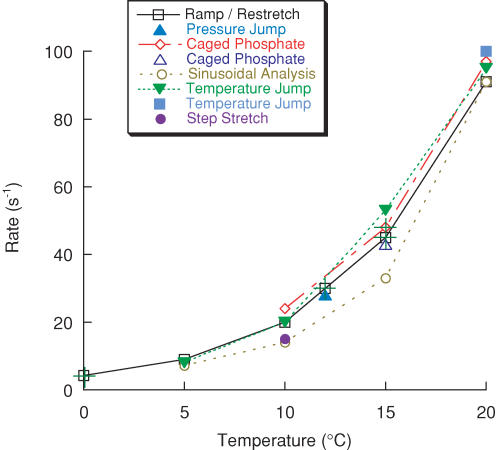
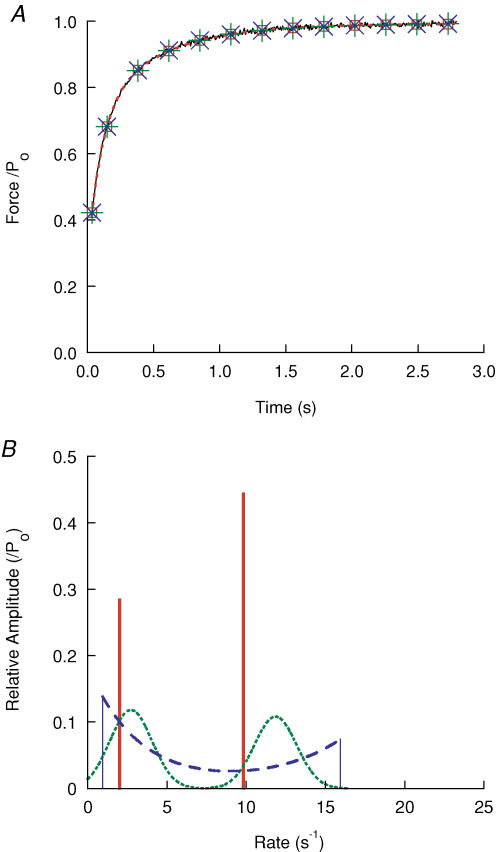
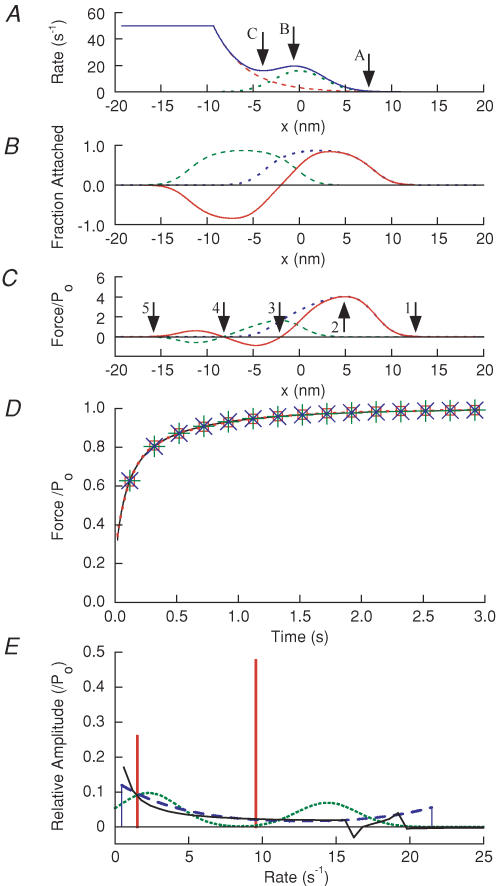
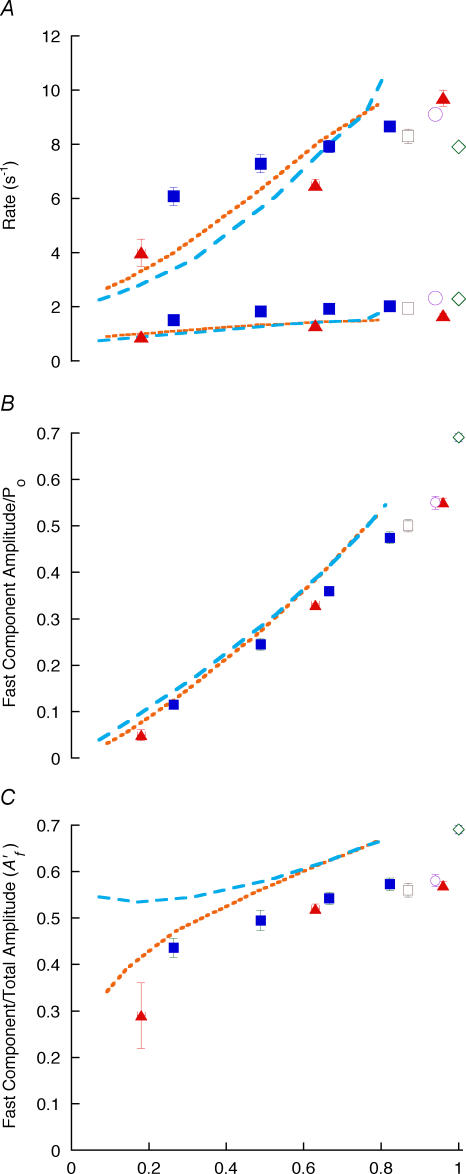
Redevelopment of isometric force following shortening of skeletal muscle is thought to result from a redistribution of cross-bridge states. We varied the initial force and cross-bridge distribution by applying various length-change protocols to active skinned single fibres from rabbit psoas muscle, and observed the effect on the slowest phase of recovery ('late recovery') that follows transient changes. In response to step releases that reduced force to near zero ( approximately 8 nm (half sarcomere)(-1)) or prolonged shortening at high velocity, late recovery was well described by two exponentials of approximately equal amplitude and rate constants of approximately 2 s(-1) and approximately 9 s(-1) at 5 degrees C. When a large restretch was applied at the end of rapid shortening, recovery was accelerated by (1) the introduction of a slow falling component that truncated the rise in force, and (2) a relative increase in the contribution of the fast exponential component. The rate of the slow fall was similar to that observed after a small isometric step stretch, with a rate of 0.4-0.8 s(-1), and its effects could be reversed by reducing force to near zero immediately after the stretch. Force at the start of late recovery was varied in a series of shortening steps or ramps in order to probe the effect of cross-bridge strain on force redevelopment. The rate constants of the two components fell by 40-50% as initial force was raised to 75-80% of steady isometric force. As initial force increased, the relative contribution of the fast component decreased, and this was associated with a length constant of about 2 nm. The results are consistent with a two-state strain-dependent cross-bridge model. In the model there is a continuous distribution of recovery rate constants, but two-exponential fits show that the fast component results from cross-bridges initially at moderate positive strain and the slow component from cross-bridges at high positive strain.
Figures
Similar articles
-
Force transients and minimum cross-bridge models in muscular contraction.J Muscle Res Cell Motil. 2007;28(7-8):371-95. doi: 10.1007/s10974-008-9131-3. Epub 2008 Apr 19. J Muscle Res Cell Motil. 2007. PMID: 18425593 Free PMC article. Review.
-
Endothermic force generation, temperature-jump experiments and effects of increased [MgADP] in rabbit psoas muscle fibres.J Physiol. 2005 Sep 1;567(Pt 2):471-92. doi: 10.1113/jphysiol.2005.090084. Epub 2005 Jun 23. J Physiol. 2005. PMID: 15975981 Free PMC article.
-
Temperature jump induced force generation in rabbit muscle fibres gets faster with shortening and shows a biphasic dependence on velocity.J Physiol. 2010 Feb 1;588(Pt 3):479-93. doi: 10.1113/jphysiol.2009.179200. Epub 2009 Nov 30. J Physiol. 2010. PMID: 19948657 Free PMC article.
-
Force generation examined by laser temperature-jumps in shortening and lengthening mammalian (rabbit psoas) muscle fibres.J Physiol. 2007 Nov 15;585(Pt 1):263-77. doi: 10.1113/jphysiol.2007.142208. Epub 2007 Oct 4. J Physiol. 2007. PMID: 17916609 Free PMC article.
-
Force and power generating mechanism(s) in active muscle as revealed from temperature perturbation studies.J Physiol. 2010 Oct 1;588(Pt 19):3657-70. doi: 10.1113/jphysiol.2010.194001. J Physiol. 2010. PMID: 20660565 Free PMC article. Review.
Cited by
-
A new experimental model to study force depression: the Drosophila jump muscle.J Appl Physiol (1985). 2014 Jun 15;116(12):1543-50. doi: 10.1152/japplphysiol.01029.2013. Epub 2014 May 1. J Appl Physiol (1985). 2014. PMID: 24790016 Free PMC article.
-
The mechanism of spontaneous oscillatory contractions in skeletal muscle.Biophys J. 2009 May 6;96(9):3682-91. doi: 10.1016/j.bpj.2009.01.039. Biophys J. 2009. PMID: 19413973 Free PMC article.
-
Force transients and minimum cross-bridge models in muscular contraction.J Muscle Res Cell Motil. 2007;28(7-8):371-95. doi: 10.1007/s10974-008-9131-3. Epub 2008 Apr 19. J Muscle Res Cell Motil. 2007. PMID: 18425593 Free PMC article. Review.
-
The rate of tension recovery in cardiac muscle correlates with the relative residual tension prevailing after restretch.Am J Physiol Heart Circ Physiol. 2007 Apr;292(4):H2020-2. doi: 10.1152/ajpheart.00714.2006. Epub 2006 Dec 22. Am J Physiol Heart Circ Physiol. 2007. PMID: 17189351 Free PMC article.
-
Insights into the kinetics of Ca2+-regulated contraction and relaxation from myofibril studies.Pflugers Arch. 2009 Jun;458(2):337-57. doi: 10.1007/s00424-008-0630-2. Epub 2009 Jan 23. Pflugers Arch. 2009. PMID: 19165498
References
-
- Bagni MA, Cecchi G, Colomo F, Tesi C. The mechanical characteristics of the contractile machinery at different levels of activation in intact single muscle fibres of the frog. In: Sugi H, Pollack GH, editors. Molecular Mechanisms of Muscle Contraction. New York: Plenum Publishing; 1988. pp. 473–488. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials