

Arabidopsis Hsa32, a novel heat shock protein, is essential for acquired thermotolerance during long recovery after acclimation
- PMID: 16500991
- PMCID: PMC1435801
- DOI: 10.1104/pp.105.074898
Arabidopsis Hsa32, a novel heat shock protein, is essential for acquired thermotolerance during long recovery after acclimation
Abstract
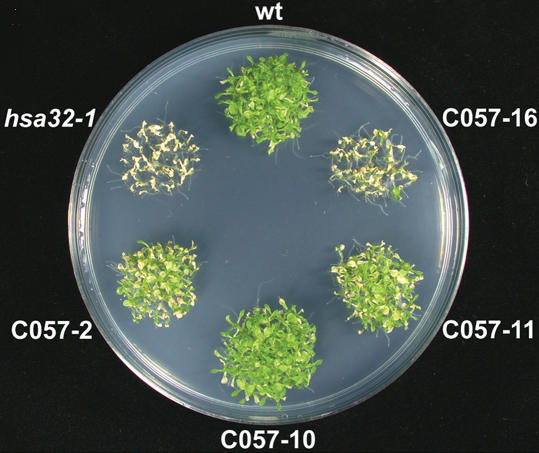
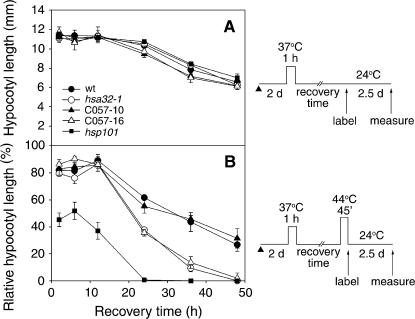
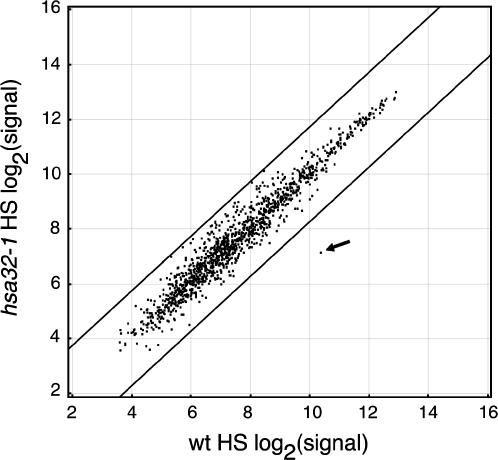
Plants and animals share similar mechanisms in the heat shock (HS) response, such as synthesis of the conserved HS proteins (Hsps). However, because plants are confined to a growing environment, in general they require unique features to cope with heat stress. Here, we report on the analysis of the function of a novel Hsp, heat-stress-associated 32-kD protein (Hsa32), which is highly conserved in land plants but absent in most other organisms. The gene responds to HS at the transcriptional level in moss (Physcomitrella patens), Arabidopsis (Arabidopsis thaliana), and rice (Oryza sativa). Like other Hsps, Hsa32 protein accumulates greatly in Arabidopsis seedlings after HS treatment. Disruption of Hsa32 by T-DNA insertion does not affect growth and development under normal conditions. However, the acquired thermotolerance in the knockout line was compromised following a long recovery period (>24 h) after acclimation HS treatment, when a severe HS challenge killed the mutant but not the wild-type plants, but no significant difference was observed if they were challenged within a short recovery period. Quantitative hypocotyl elongation assay also revealed that thermotolerance decayed faster in the absence of Hsa32 after a long recovery. Similar results were obtained in Arabidopsis transgenic plants with Hsa32 expression suppressed by RNA interference. Microarray analysis of the knockout mutant indicates that only the expression of Hsa32 was significantly altered in HS response. Taken together, our results suggest that Hsa32 is required not for induction but rather maintenance of acquired thermotolerance, a feature that could be important to plants.
Figures
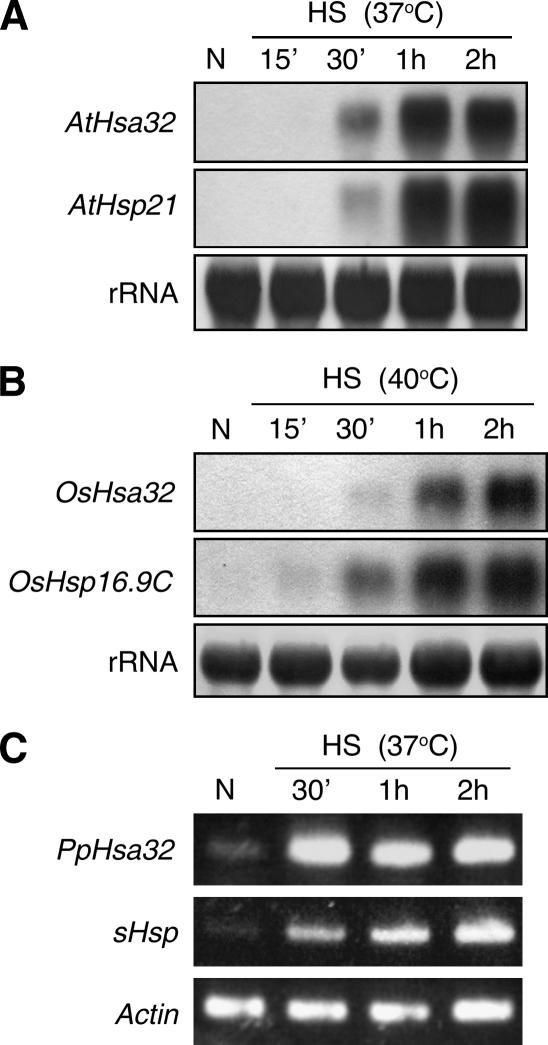
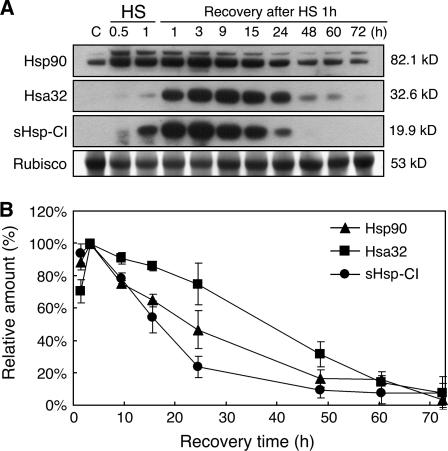
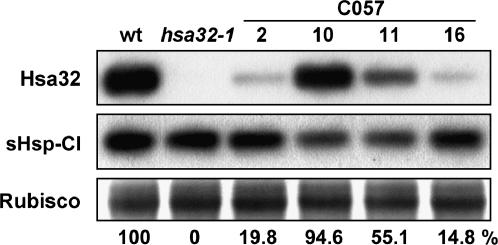
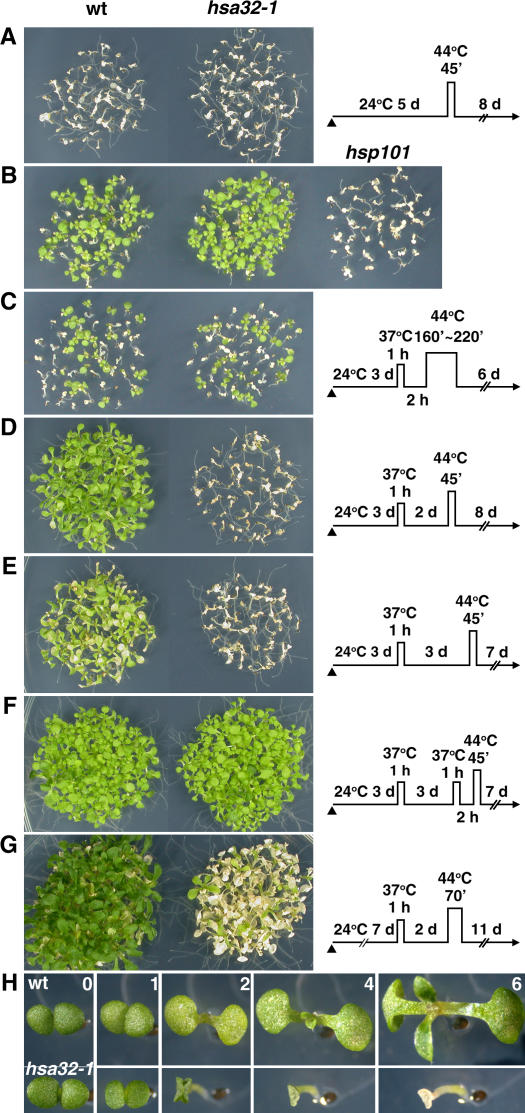
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Burke JJ (2001) Identification of genetic diversity and mutations in higher plant acquired thermotolerance. Physiol Plant 112: 167–170
-
- Busch W, Wunderlich M, Schoffl F (2005) Identification of novel heat shock factor-dependent genes and biochemical pathways in Arabidopsis thaliana. Plant J 41: 1–14 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
