

Calmodulin interacts with and regulates the RNA-binding activity of an Arabidopsis polyadenylation factor subunit
- PMID: 16500995
- PMCID: PMC1459842
- DOI: 10.1104/pp.105.070672
Calmodulin interacts with and regulates the RNA-binding activity of an Arabidopsis polyadenylation factor subunit
Abstract
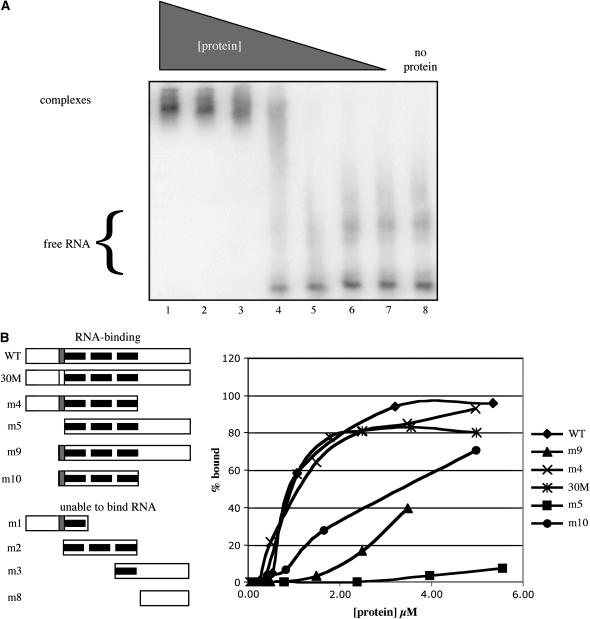
The Arabidopsis (Arabidopsis thaliana) gene that encodes the probable ortholog of the 30-kD subunit of the mammalian cleavage and polyadenylation specificity factor (CPSF) is a complex one, encoding small (approximately 28 kD) and large (approximately 68 kD) polypeptides. The small polypeptide (AtCPSF30) corresponds to CPSF30 and is the focus of this study. Recombinant AtCPSF30 was purified from Escherichia coli and found to possess RNA-binding activity. Mutational analysis indicated that an evolutionarily conserved central core of AtCPSF30 is involved in RNA binding, but that RNA binding also requires a short sequence adjacent to the N terminus of the central core. AtCPSF30 was found to bind calmodulin, and calmodulin inhibited the RNA-binding activity of the protein in a calcium-dependent manner. Mutational analysis showed that a small part of the protein, again adjacent to the N terminus of the conserved core, is responsible for calmodulin binding; point mutations in this region abolished both binding to and inhibition of RNA binding by calmodulin. Interestingly, AtCPSF30 was capable of self-interactions. This property also mapped to the central conserved core of the protein. However, calmodulin had no discernible effect on the self-association. These results show that the central portion of AtCPSF30 is involved in a number of important functions, and they raise interesting possibilities for both the interplay between splicing and polyadenylation and the regulation of these processes by stimuli that act through calmodulin.
Figures








Similar articles
-
Redox and heavy metal effects on the biochemical activities of an Arabidopsis polyadenylation factor subunit.Arch Biochem Biophys. 2008 May 1;473(1):88-95. doi: 10.1016/j.abb.2008.02.027. Epub 2008 Mar 4. Arch Biochem Biophys. 2008. PMID: 18331819
-
A novel endonuclease activity associated with the Arabidopsis ortholog of the 30-kDa subunit of cleavage and polyadenylation specificity factor.Nucleic Acids Res. 2007;35(13):4453-63. doi: 10.1093/nar/gkm457. Epub 2007 Jun 18. Nucleic Acids Res. 2007. PMID: 17576667 Free PMC article.
-
A disulfide linkage in a CCCH zinc finger motif of an Arabidopsis CPSF30 ortholog.FEBS Lett. 2010 Nov 5;584(21):4408-12. doi: 10.1016/j.febslet.2010.09.043. Epub 2010 Oct 1. FEBS Lett. 2010. PMID: 20888817
-
CPSF30 at the Interface of Alternative Polyadenylation and Cellular Signaling in Plants.Biomolecules. 2015 Jun 8;5(2):1151-68. doi: 10.3390/biom5021151. Biomolecules. 2015. PMID: 26061761 Free PMC article. Review.
-
The Arabidopsis polyadenylation factor subunit CPSF30 as conceptual link between mRNA polyadenylation and cellular signaling.Curr Opin Plant Biol. 2014 Oct;21:128-132. doi: 10.1016/j.pbi.2014.07.002. Epub 2014 Aug 9. Curr Opin Plant Biol. 2014. PMID: 25104048 Review.
Cited by
-
Plant calmodulins and calmodulin-related proteins: multifaceted relays to decode calcium signals.Plant Signal Behav. 2006 May;1(3):96-104. doi: 10.4161/psb.1.3.2998. Plant Signal Behav. 2006. PMID: 19521489 Free PMC article.
-
Plant terminators: the unsung heroes of gene expression.J Exp Bot. 2023 Apr 9;74(7):2239-2250. doi: 10.1093/jxb/erac467. J Exp Bot. 2023. PMID: 36477559 Free PMC article. Review.
-
Putative molecular mechanisms underlying tandem CCCH zinc finger protein mediated plant growth, stress, and gene expression responses.Plant Signal Behav. 2011 May;6(5):647-51. doi: 10.4161/psb.6.5.15105. Plant Signal Behav. 2011. PMID: 21795857 Free PMC article. Review.
-
FIP1 Plays an Important Role in Nitrate Signaling and Regulates CIPK8 and CIPK23 Expression in Arabidopsis.Front Plant Sci. 2018 May 4;9:593. doi: 10.3389/fpls.2018.00593. eCollection 2018. Front Plant Sci. 2018. PMID: 29780398 Free PMC article.
-
Cleavage and polyadenylation: Ending the message expands gene regulation.RNA Biol. 2017 Jul 3;14(7):865-890. doi: 10.1080/15476286.2017.1306171. Epub 2017 Apr 28. RNA Biol. 2017. PMID: 28453393 Free PMC article.
References
-
- Addepalli B, Meeks LR, Forbes KP, Hunt AG (2004) Novel alternative splicing of mRNAs encoding poly(A) polymerases in Arabidopsis. Biochim Biophys Acta 1679: 117–128 - PubMed
-
- Amasino RM (2003) Flowering time: a pathway that begins at the 3′ end. Curr Biol 13: 670–672 - PubMed
-
- Barabino SM, Hubner W, Jenny A, Minvielle-Sebastia L, Keller W (1997) The 30-kD subunit of mammalian cleavage and polyadenylation specificity factor and its yeast homolog are RNA-binding zinc finger proteins. Genes Dev 11: 1703–1716 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
