

Projection structure of the human copper transporter CTR1 at 6-A resolution reveals a compact trimer with a novel channel-like architecture
- PMID: 16501047
- PMCID: PMC1450133
- DOI: 10.1073/pnas.0509929103
Projection structure of the human copper transporter CTR1 at 6-A resolution reveals a compact trimer with a novel channel-like architecture
Abstract
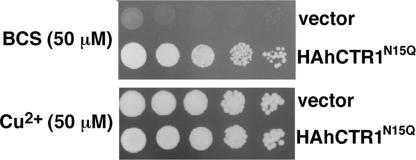
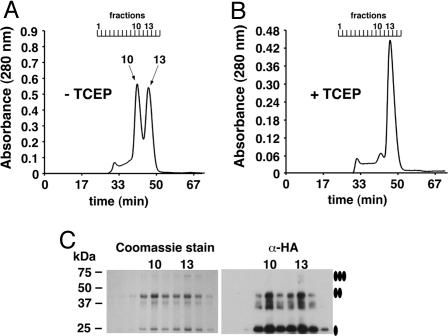
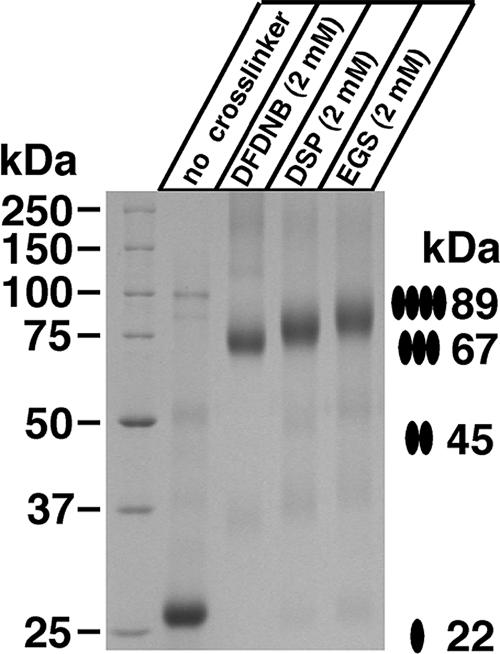
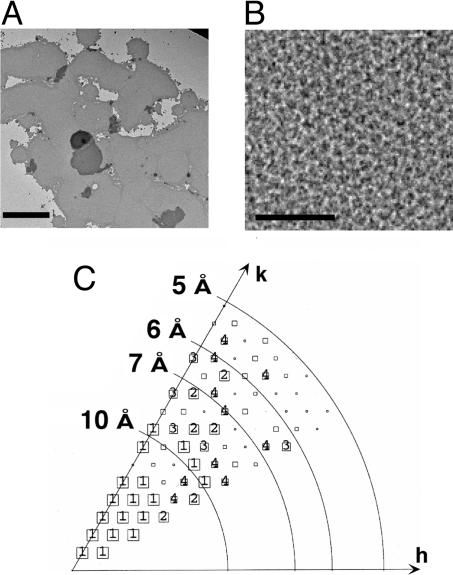
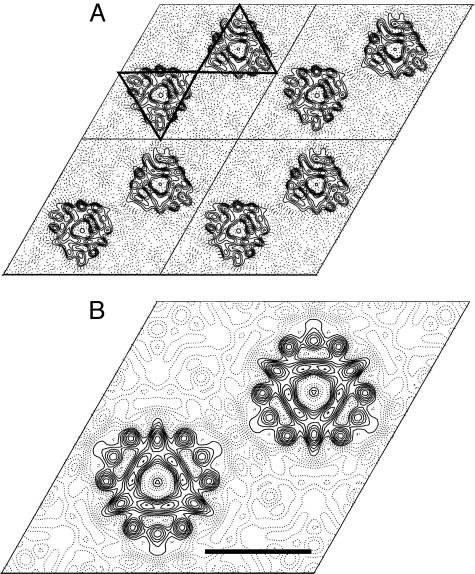
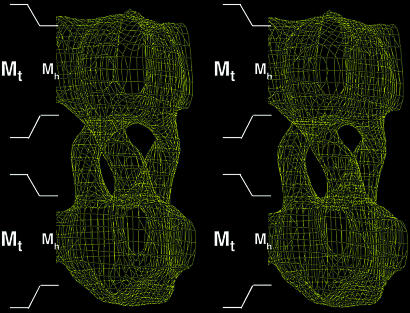
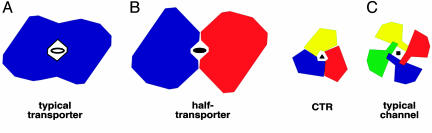
Human CTR1 is a high-affinity copper transporter that also mediates the uptake of the anticancer drug cisplatin by largely unknown transport mechanisms. Here we report the 6-A projection structure obtained for human CTR1 by using electron crystallography of 2D protein crystals in a native phospholipid bilayer. The projection of CTR1 reveals a symmetrical trimer that is <40 A wide. Notably, the center threefold axis of each trimer forms a region of very low electron density likely to be involved in copper translocation. The formation of a putative pore for metal ions at the interface of three identical subunits deviates from the structural design of typical primary and secondary active transporters and reveals that copper uptake transporters have a novel architecture that is structurally more closely related to channel proteins.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
