

RNase L plays a role in the antiviral response to West Nile virus
- PMID: 16501108
- PMCID: PMC1395436
- DOI: 10.1128/JVI.80.6.2987-2999.2006
RNase L plays a role in the antiviral response to West Nile virus
Abstract
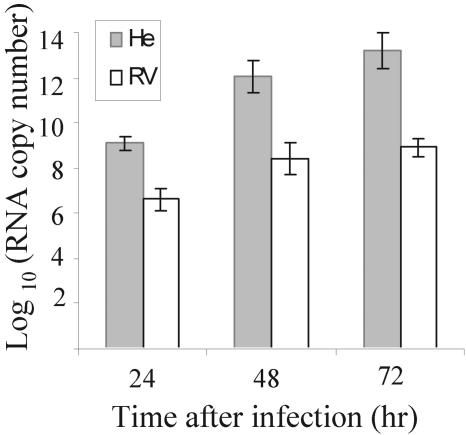
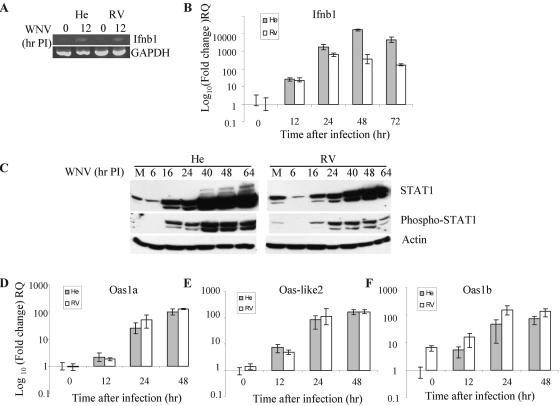
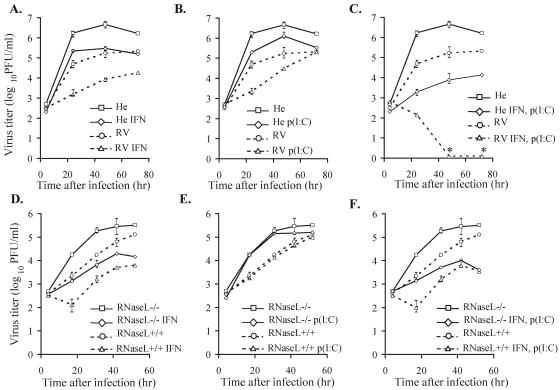
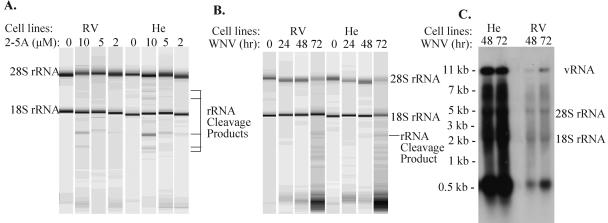
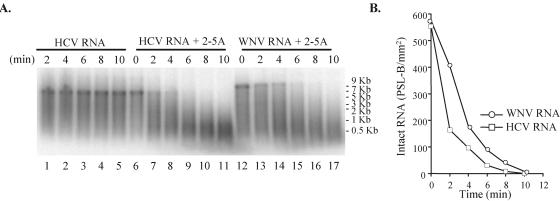
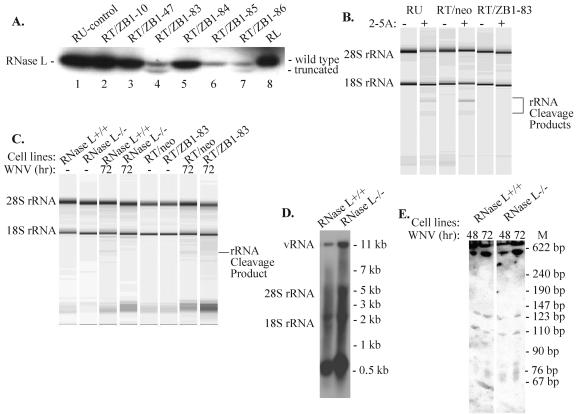
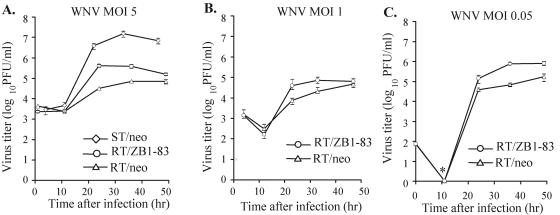
Alleles at the Flv locus determine disease outcome after a flavivirus infection in mice. Although comparable numbers of congenic resistant and susceptible mouse embryo fibroblasts (MEFs) are infected by the flavivirus West Nile virus (WNV), resistant MEFs produce approximately 100- to 150-fold lower titers than susceptible ones and flavivirus titers in the brains of resistant and susceptible animals can differ by >10,000-fold. The Flv locus was previously identified as the 2'-5' oligoadenylate synthetase 1b (Oas1b) gene. Oas gene expression is up-regulated by interferon (IFN), and after activation by double-stranded RNA, some mouse synthetases produce 2-5A, which activates latent RNase L to degrade viral and cellular RNAs. To determine whether the lower levels of intracellular flavivirus genomic RNA from resistant mice detected in cells at all times after infection were mediated by RNase L, RNase L activity levels in congenic resistant and susceptible cells were compared. Similar moderate levels of RNase L activation by transfected 2-5A were observed in both types of uninfected cells. After WNV infection, the mRNAs of IFN-beta and three Oas genes were up-regulated to similar levels in both types of cells. However, significant levels of RNase L activity were not detected until 72 h after WNV infection and the patterns of viral RNA cleavage products generated were similar in both types of cells. When RNase L activity was down-regulated in resistant cells via stable expression of a dominant negative RNase L mutant, approximately 5- to 10-times-higher yields of WNV were produced. Similarly, about approximately 5- to 10-times-higher virus yields were produced by susceptible C57BL/6 RNase L-/- cells compared to RNase L+/+ cells that were either left untreated or pretreated with IFN and/or poly(I) . poly(C). The data indicate that WNV genomic RNA is susceptible to RNase L cleavage and that RNase L plays a role in the cellular antiviral response to flaviviruses. The results suggest that RNase L activation is not a major component of the Oas1b-mediated flavivirus resistance phenotype.
Figures
Similar articles
-
RNase L Antiviral Activity Is Not a Critical Component of the Oas1b-Mediated Flavivirus Resistance Phenotype.J Virol. 2019 Oct 29;93(22):e00946-19. doi: 10.1128/JVI.00946-19. Print 2019 Nov 15. J Virol. 2019. PMID: 31462564 Free PMC article.
-
The 2',5'-oligoadenylate synthetase 1b is a potent inhibitor of West Nile virus replication inside infected cells.J Biol Chem. 2006 Feb 24;281(8):4624-37. doi: 10.1074/jbc.M508649200. Epub 2005 Dec 21. J Biol Chem. 2006. PMID: 16371364
-
The Flvr-encoded murine oligoadenylate synthetase 1b (Oas1b) suppresses 2-5A synthesis in intact cells.Virology. 2011 Jan 20;409(2):262-70. doi: 10.1016/j.virol.2010.10.016. Epub 2010 Nov 5. Virology. 2011. PMID: 21056894 Free PMC article.
-
The 2-5A system in viral infection and apoptosis.Biomed Pharmacother. 1998;52(9):386-90. doi: 10.1016/s0753-3322(99)80006-7. Biomed Pharmacother. 1998. PMID: 9856285 Review.
-
The Many Faces of Oligoadenylate Synthetases.J Interferon Cytokine Res. 2023 Nov;43(11):487-494. doi: 10.1089/jir.2023.0098. Epub 2023 Sep 25. J Interferon Cytokine Res. 2023. PMID: 37751211 Free PMC article. Review.
Cited by
-
West Nile virus infection and immunity.Nat Rev Microbiol. 2013 Feb;11(2):115-28. doi: 10.1038/nrmicro2950. Nat Rev Microbiol. 2013. PMID: 23321534 Review.
-
West Nile virus noncoding subgenomic RNA contributes to viral evasion of the type I interferon-mediated antiviral response.J Virol. 2012 May;86(10):5708-18. doi: 10.1128/JVI.00207-12. Epub 2012 Feb 29. J Virol. 2012. PMID: 22379089 Free PMC article.
-
Association between HLA class I and class II alleles and the outcome of West Nile virus infection: an exploratory study.PLoS One. 2011;6(8):e22948. doi: 10.1371/journal.pone.0022948. Epub 2011 Aug 1. PLoS One. 2011. PMID: 21829673 Free PMC article.
-
Replication-Dependent Biogenesis of Turnip Crinkle Virus Long Noncoding RNAs.J Virol. 2021 Aug 25;95(18):e0016921. doi: 10.1128/JVI.00169-21. Epub 2021 Aug 25. J Virol. 2021. PMID: 34160262 Free PMC article.
-
Antagonism of the interferon-induced OAS-RNase L pathway by murine coronavirus ns2 protein is required for virus replication and liver pathology.Cell Host Microbe. 2012 Jun 14;11(6):607-16. doi: 10.1016/j.chom.2012.04.011. Cell Host Microbe. 2012. PMID: 22704621 Free PMC article.
References
-
- Al-khatib, K., B. R. Williams, R. Silverman, W. P. Halford, and D. J. Carr. 2003. The murine double-stranded RNA-dependent protein kinase PKR and murine 2′,5′-oligoadenylate synthetase-dependent RNase L are required for IFN-β-mediated resistance against herpes simplex virus type 1 in primary trigeminal ganglion culture. Virology 313:126-135. - PubMed
-
- Bhatt, P. N., E. A. Johnson, A. L. Smith, and R. O. Jacoby. 1981. Genetic resistance to lethal flavivirus encephalitis. III. Replication of Banzi virus in vitro and in vivo in tissues of congenic susceptible and resistant mice. Arch. Virol. 69:273-286. - PubMed
-
- Biron, C., and G. C. Sen. 2001. Interferons and other cytokines, p. 321-351. In D. M. Knipe, P. M. Howley, D. E. Griffin, R. A. Lamb, M. A. Martin, B. Roizman, and S. E. Straus (ed.), Fields virology, 4th edition. Lippincott-Raven Publishers, Philadelphia, Pa.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
