

The early history of the ubiquitin field
- PMID: 16501229
- PMCID: PMC2249785
- DOI: 10.1110/ps.052012306
The early history of the ubiquitin field
Abstract
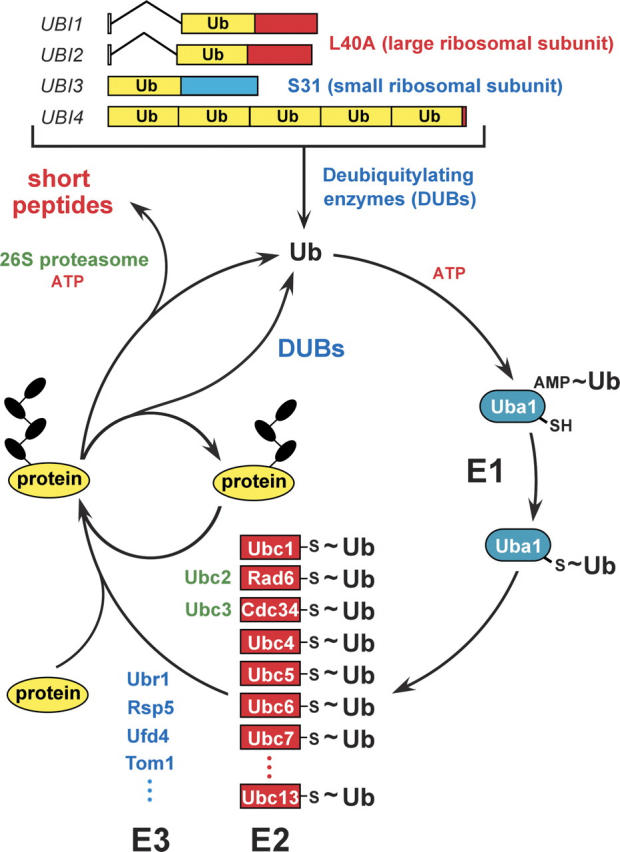
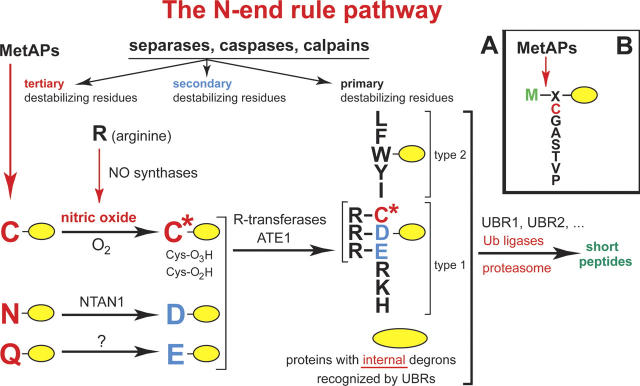
This is a personal account of the early history of ubiquitin research, by one of its protagonists. The field of ubiquitin and regulated protein degradation was created in the 1980s, largely through the complementary discoveries by the laboratory of A. Hershko (Technion, Haifa, Israel) and by my laboratory, then at MIT (Cambridge, MA). I describe the elegant insights by Hershko and his colleagues that yielded the initial understanding of ubiquitin conjugation and ubiquitin-mediated proteolysis in cell extracts, including the identification of E1, E2, and E3 enzymes. These advances were followed by a set of interconnected discoveries in my laboratory that revealed the biology of the ubiquitin system, i.e., its necessity for the protein degradation in vivo, its specific physiological functions (in the cell cycle, DNA repair, protein synthesis, transcriptional regulation, and stress responses), the source of its selectivity (specific degradation signals in short-lived proteins), and its key mechanistic attributes, such as the polyubiquitin chain and the subunit selectivity of protein degradation. The above biological (function-based) insights produced the main discovery of the physiological regulation by intracellular protein degradation. These advances caused the enormous expansion of the ubiquitin field in the 1990s. Together with the initial discovery of ubiquitin-mediated proteolysis by Hershko and coworkers, our biological discoveries in the 1980s led to a radically changed understanding of the logic of intracellular circuits, as it became clear that the control through regulated protein degradation rivals, and often surpasses in significance, the classical regulation through transcription and translation.
Figures
References
-
- Bachmair, A. and Varshavsky, A. 1989. The degradation signal in a short-lived protein. Cell 56: 1019–1032. - PubMed
-
- Bachmair, A., Finley, D., and Varshavsky, A. 1986. In vivo half-life of a protein is a function of its amino-terminal residue. Science 234: 179–186. - PubMed
-
- Baumeister, W., Walz, J., Zühl, F., and Seemüller, E. 1998. The proteasome: Paradigm of a self-compartmentalizing protease. Cell 92: 367–380. - PubMed
-
- Chau, V., Tobias, J.W., Bachmair, A., Marriott, D., Ecker, D.J., Gonda, D.K., and Varshavsky, A. 1989. A multiubiquitin chain is confined to specific lysine in a targeted short-lived protein. Science 243: 1576–1583. - PubMed
Publication types
MeSH terms
Substances
Personal name as subject
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
