

Modulation of reactivity and conformation within the T-quaternary state of human hemoglobin: the combined use of mutagenesis and sol-gel encapsulation
- PMID: 16503637
- PMCID: PMC3558951
- DOI: 10.1021/bi050010i
Modulation of reactivity and conformation within the T-quaternary state of human hemoglobin: the combined use of mutagenesis and sol-gel encapsulation
Abstract
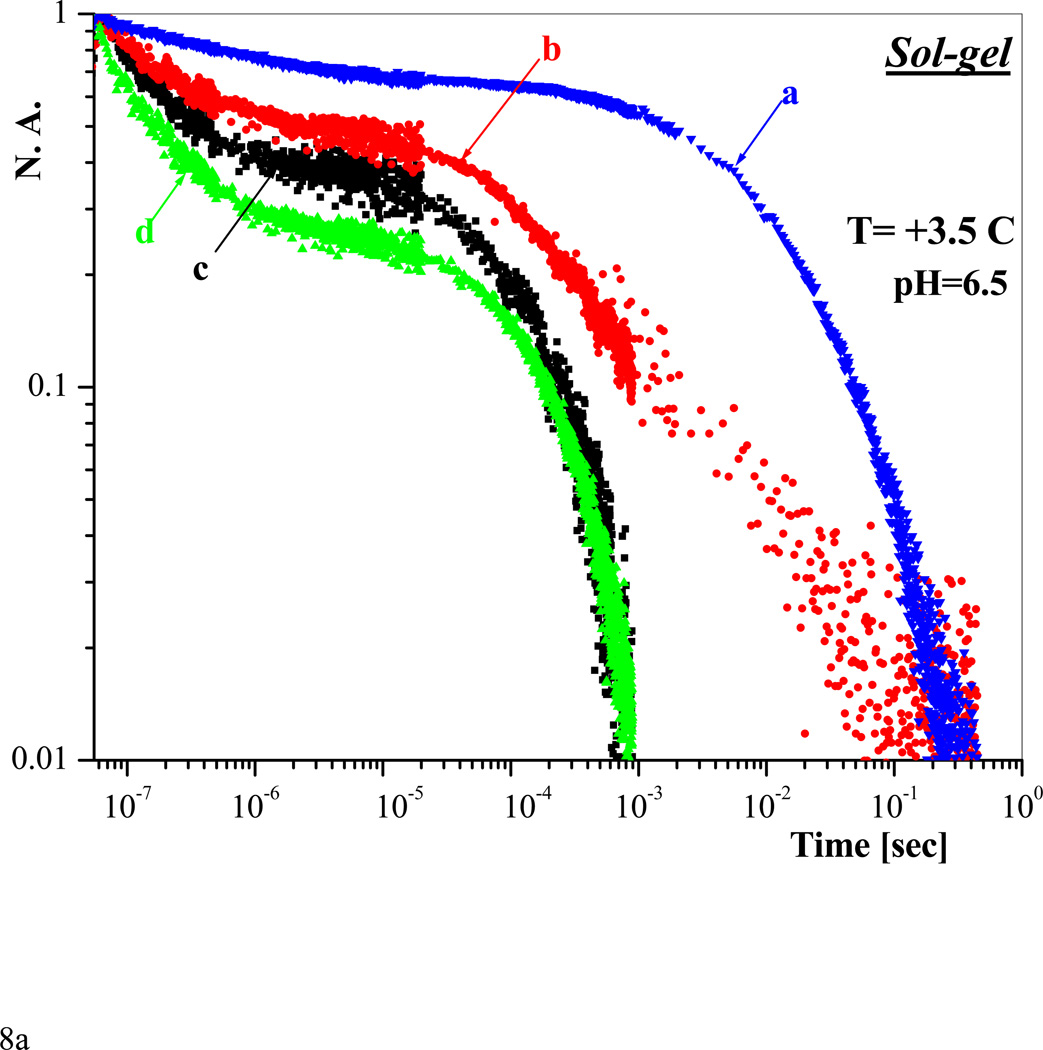
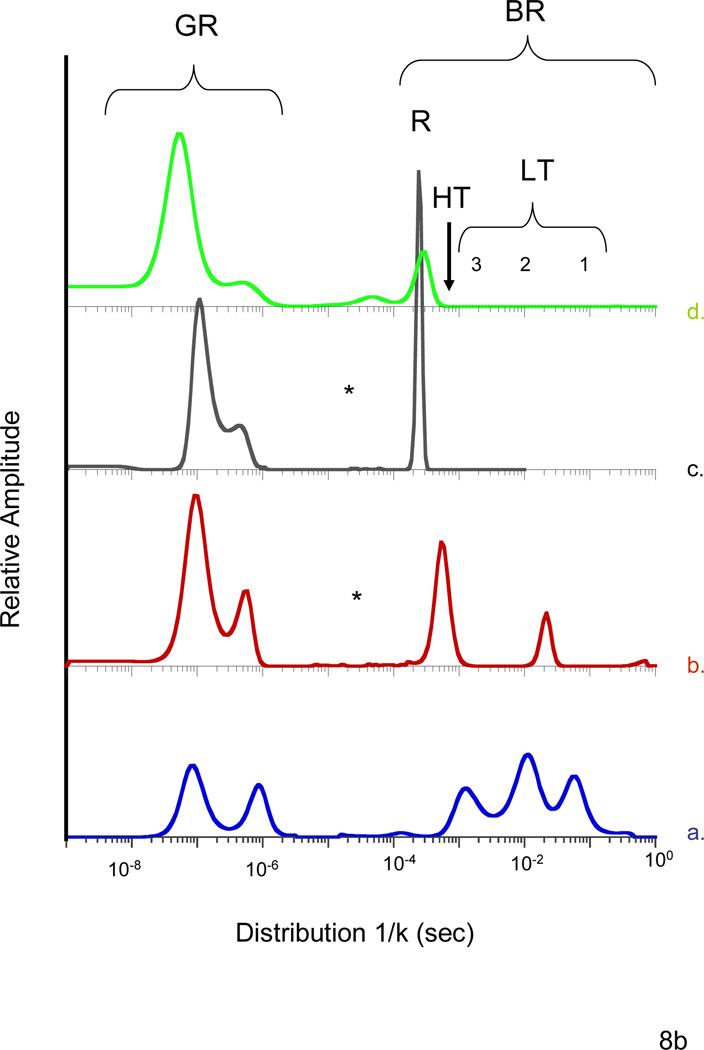
A range of conformationally distinct functional states within the T quaternary state of hemoglobin are accessed and probed using a combination of mutagenesis and sol-gel encapsulation that greatly slow or eliminate the T --> R transition. Visible and UV resonance Raman spectroscopy are used to probe the proximal strain at the heme and the status of the alpha(1)beta(2) interface, respectively, whereas CO geminate and bimolecular recombination traces in conjunction with MEM (maximum entropy method) analysis of kinetic populations are used to identify functionally distinct T-state populations. The mutants used in this study are Hb(Nbeta102A) and the alpha99-alpha99 cross-linked derivative of Hb(Wbeta37E). The former mutant, which binds oxygen noncooperatively with very low affinity, is used to access low-affinity ligated T-state conformations, whereas the latter mutant is used to access the high-affinity end of the distribution of T-state conformations. A pattern emerges within the T state in which ligand reactivity increases as both the proximal strain and the alpha(1)beta(2) interface interactions are progressively lessened after ligand binding to the deoxy T-state species. The ligation and effector-dependent interplay between the heme environment and the stability of the Trp beta37 cluster in the hinge region of the alpha(1)beta(2) interface appears to determine the distribution of the ligated T-state species generated upon ligand binding. A qualitative model is presented, suggesting that different T quaternary structures modulate the stability of different alphabeta dimer conformations within the tetramer.
Figures
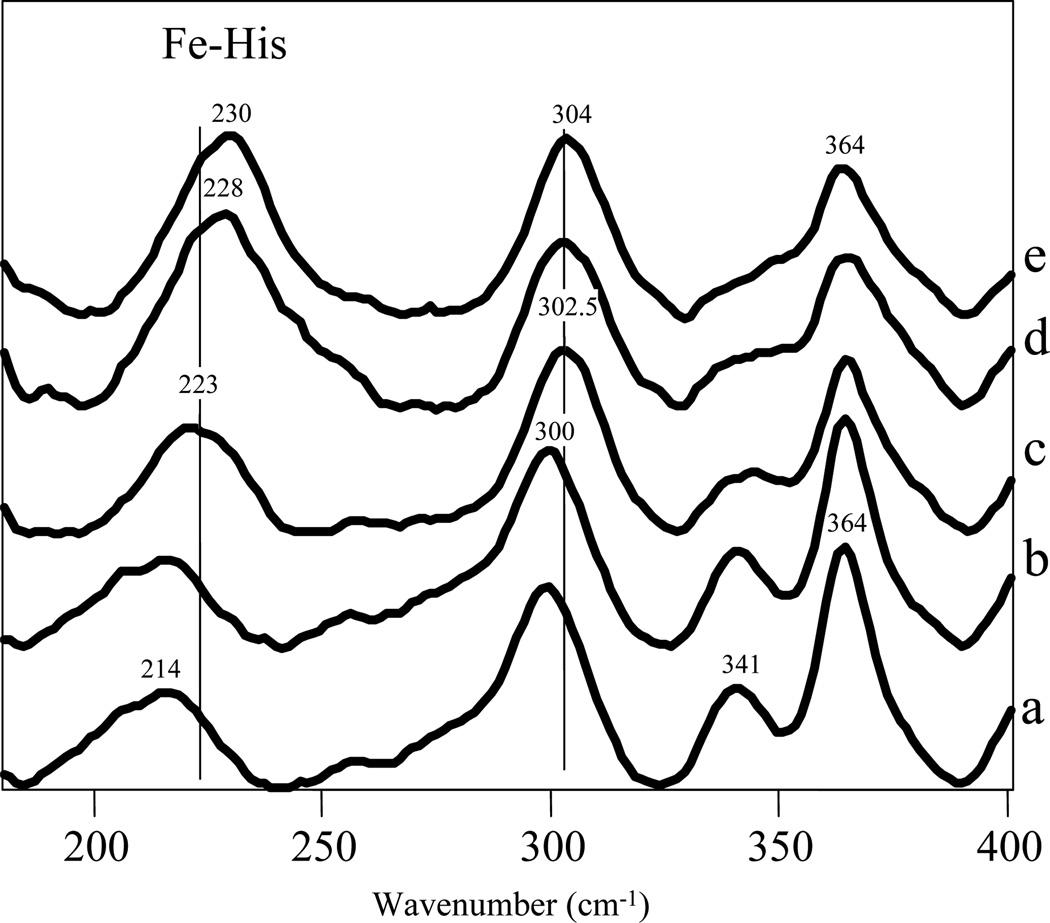

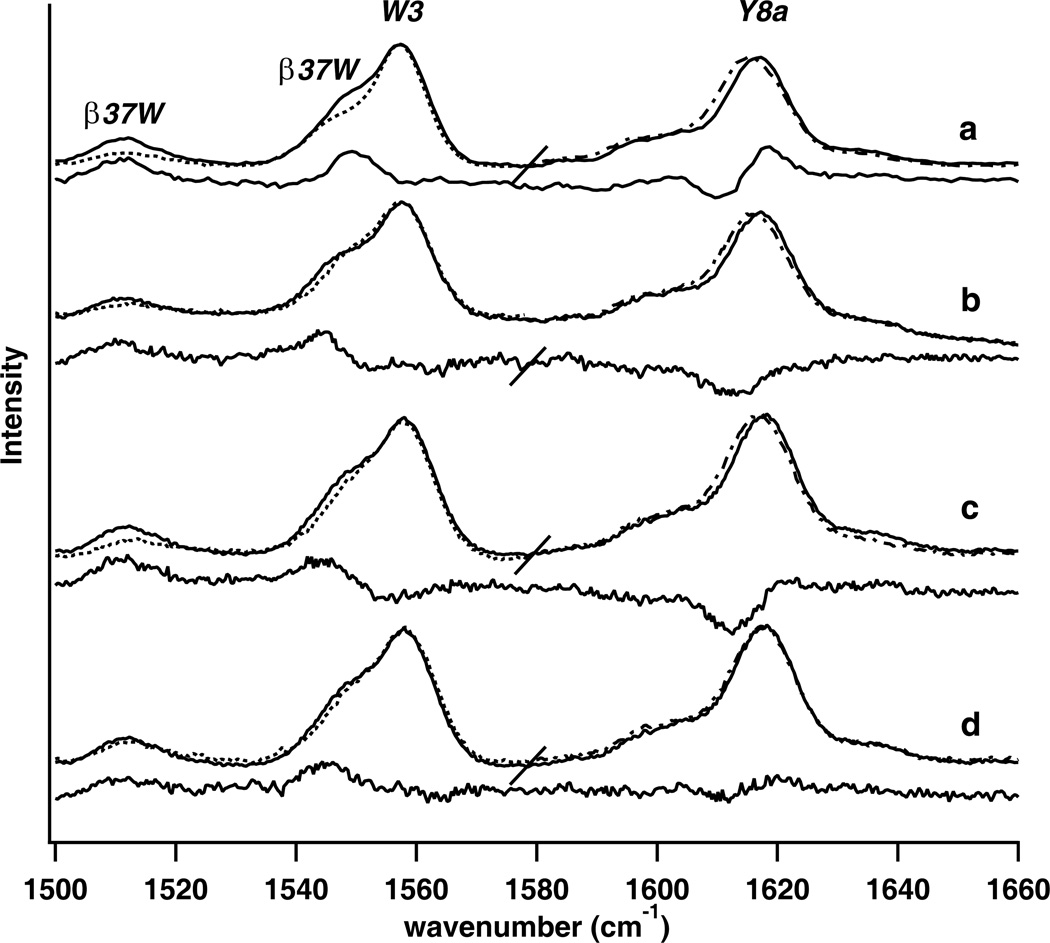



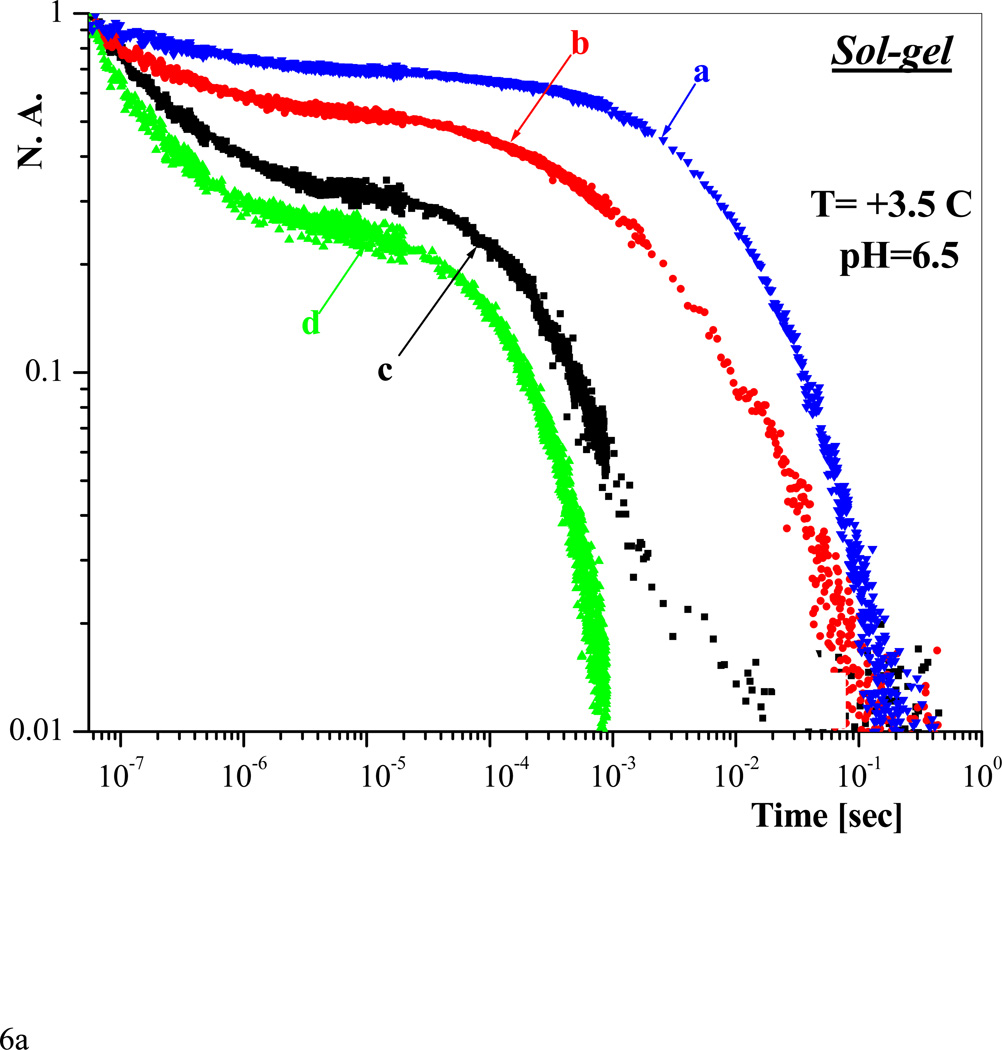


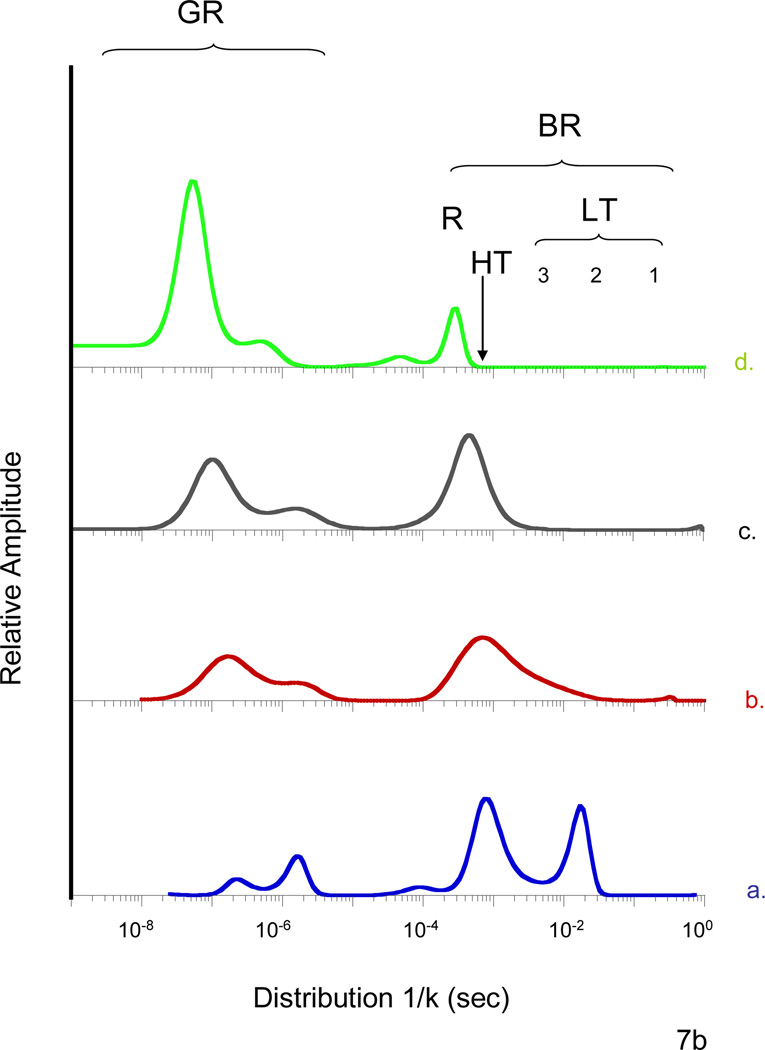

Similar articles
-
Spectroscopic and functional characterization of T state hemoglobin conformations encapsulated in silica gels.Biochemistry. 2004 Nov 2;43(43):13674-82. doi: 10.1021/bi048531d. Biochemistry. 2004. PMID: 15504030
-
A possible allosteric communication pathway identified through a resonance Raman study of four beta37 mutants of human hemoglobin A.Biochemistry. 1998 Mar 31;37(13):4346-57. doi: 10.1021/bi9708693. Biochemistry. 1998. PMID: 9521755
-
Functional and spectroscopic characterization of half-liganded iron-zinc hybrid hemoglobin: evidence for conformational plasticity within the T state.Biochemistry. 2003 Jul 15;42(27):8272-88. doi: 10.1021/bi020648j. Biochemistry. 2003. PMID: 12846576
-
Transduction of binding energy into hemoglobin cooperativity.Trends Biochem Sci. 1993 Oct;18(10):385-90. doi: 10.1016/0968-0004(93)90095-5. Trends Biochem Sci. 1993. PMID: 8256288 Review.
-
The functional role of the hemoglobin-water interface.Mol Aspects Med. 2022 Apr;84:101042. doi: 10.1016/j.mam.2021.101042. Epub 2021 Oct 29. Mol Aspects Med. 2022. PMID: 34756740 Review.
Cited by
-
Generating S-nitrosothiols from hemoglobin: mechanisms, conformational dependence, and physiological relevance.J Biol Chem. 2013 Aug 2;288(31):22408-25. doi: 10.1074/jbc.M113.482679. Epub 2013 Jun 17. J Biol Chem. 2013. PMID: 23775069 Free PMC article.
-
Protein dynamics from time resolved UV Raman spectroscopy.Curr Opin Struct Biol. 2008 Oct;18(5):623-9. doi: 10.1016/j.sbi.2008.06.001. Epub 2008 Jul 19. Curr Opin Struct Biol. 2008. PMID: 18606227 Free PMC article. Review.
-
NO reactions with sol-gel and solution phase samples of the ferric nitrite derivative of HbA.Nitric Oxide. 2010 Feb 15;22(2):180-90. doi: 10.1016/j.niox.2009.11.003. Epub 2009 Nov 15. Nitric Oxide. 2010. PMID: 19919854 Free PMC article.
-
Circular dichroism spectroscopy of tertiary and quaternary conformations of human hemoglobin entrapped in wet silica gels.Protein Sci. 2006 Aug;15(8):1961-7. doi: 10.1110/ps.062272306. Epub 2006 Jul 5. Protein Sci. 2006. PMID: 16823042 Free PMC article.
-
Tertiary and quaternary allostery in tetrameric hemoglobin from Scapharca inaequivalvis.Biochemistry. 2013 Mar 26;52(12):2108-17. doi: 10.1021/bi301620x. Epub 2013 Mar 15. Biochemistry. 2013. PMID: 23458680 Free PMC article.
References
-
- Monod J, Wyman J, Changeux JP. On the Nature of Allosteric Transitions: A Plausible Model. J Mol Biol. 1965;12:88–118. - PubMed
-
- Eaton WA, Henry ER, Hofrichter J, Mozzarelli A. Is cooperative oxygen binding by hemoglobin really understood? Nat Struct Biol. 1999;6:351–358. - PubMed
-
- Perutz MF, Fermi G, Luisi B, Shaanan B, Liddington RC. Stereochemistry of cooperative mechanisms in hemoglobin. Cold Spring Harb Symp Quant Biol. 1987;52:555–565. - PubMed
-
- Perutz MF, Wilkinson AJ, Paoli M, Dodson GG. The stereochemical mechanism of the cooperative effects in hemoglobin revisited. Annu Rev Biophys Biomol Struct. 1998;27:1–34. - PubMed
-
- Shulman RG, Ogawa S, Hopfield JJ. An allosteric model of hemoglobin. Cold Spring Harb Symp Quant Biol. 1972;36:337–341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
