

The complement system plays a critical role in the development of experimental autoimmune anterior uveitis
- PMID: 16505038
- PMCID: PMC1975680
- DOI: 10.1167/iovs.05-1062
The complement system plays a critical role in the development of experimental autoimmune anterior uveitis
Abstract
Purpose: The role of complement in ocular autoimmunity was explored in a experimental autoimmune anterior uveitis (EAAU) animal model.
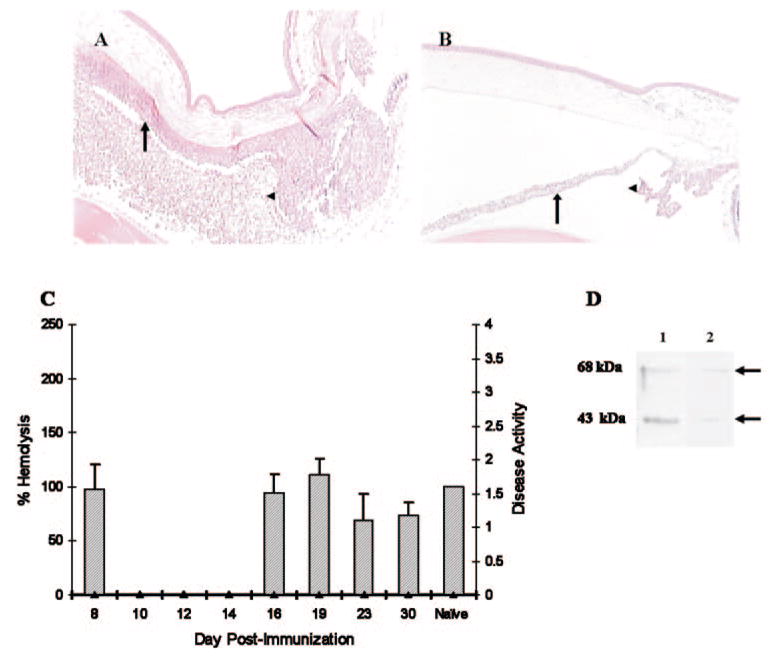
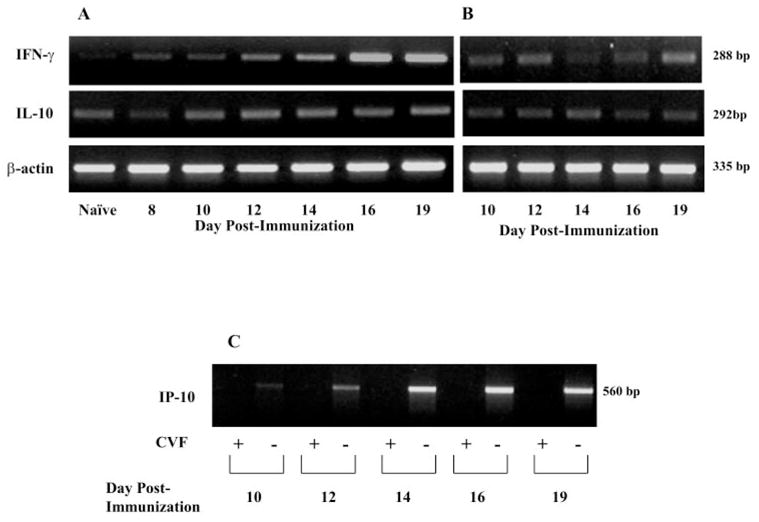
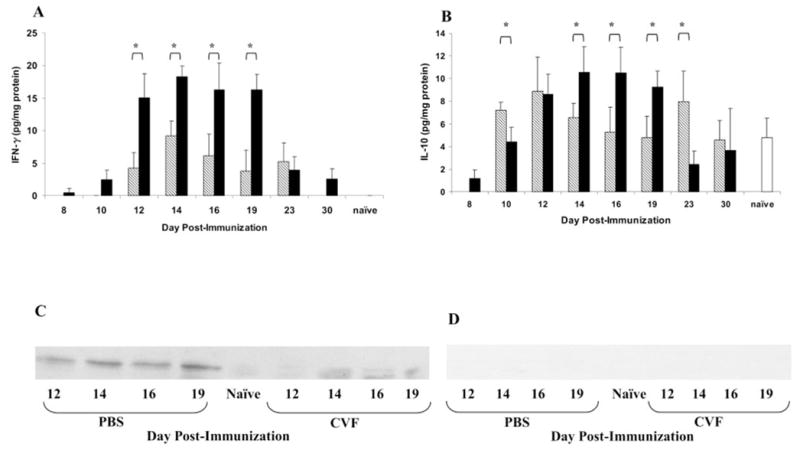
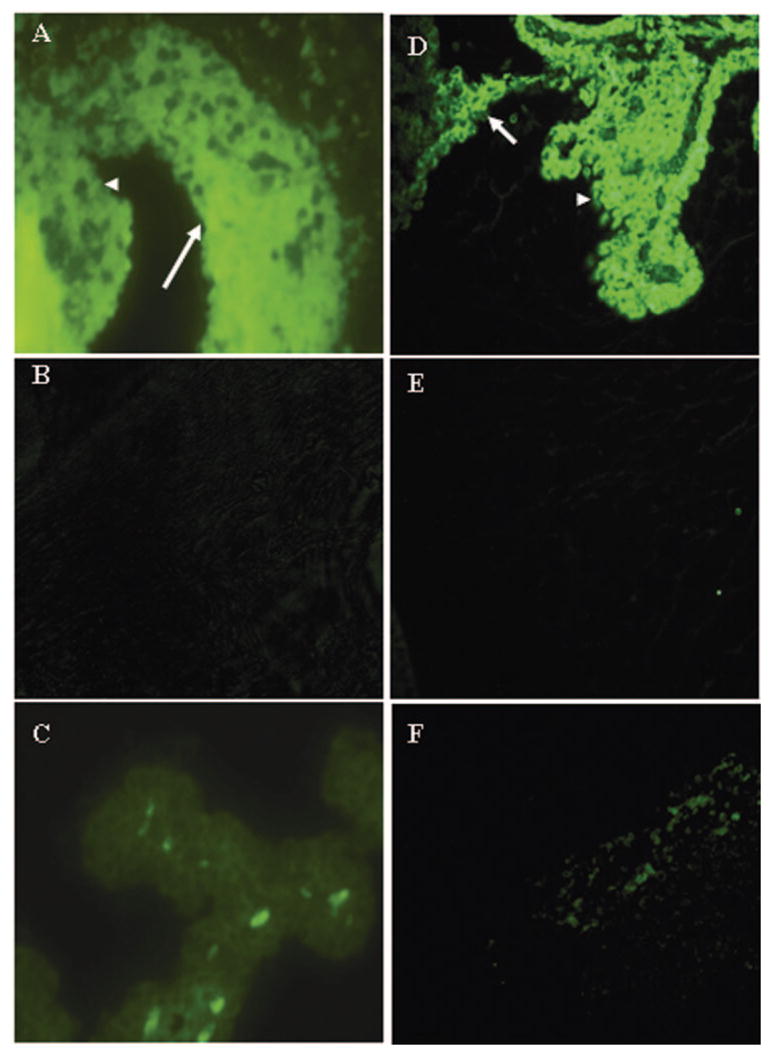
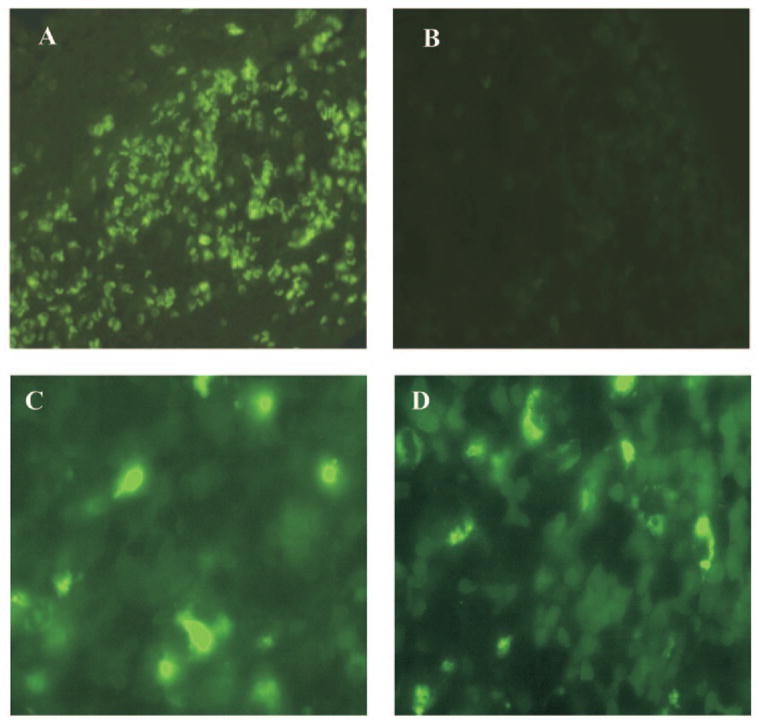
Methods: EAAU was induced in Lewis rats by immunization with bovine melanin-associated antigen. Complement activation in the eye was monitored by Western blot for iC3b. The importance of complement to the development of EAAU was studied by comparing the course of intraocular inflammation in normal Lewis rats (complement-sufficient) with cobra venom factor-treated rats (complement-depleted). Eyes were harvested from both complement-sufficient and complement-depleted rats for mRNA and protein analysis for IFN-gamma, IL-10, and interferon-inducible protein (IP)-10. Intracellular adhesion molecule (ICAM)-1 and leukocyte-endothelial cell adhesion molecule (LECAM)-1 were detected by immunofluorescent staining. OX-42 was used to investigate the importance of iC3b and CR3 interaction in EAAU.
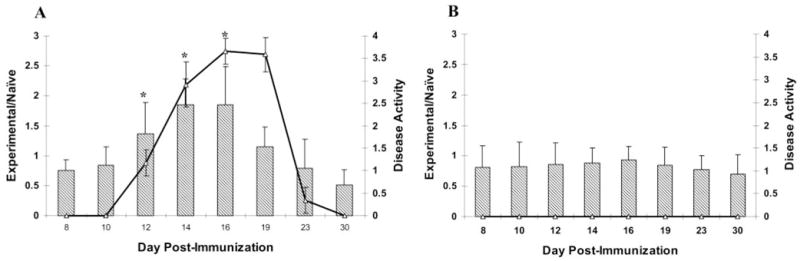
Results: There was a correlation between ocular complement activation and disease progression in EAAU. The incidence, duration, and severity of disease were dramatically reduced after active immunization in complement-depleted rats. Complement depletion also completely suppressed adoptive transfer EAAU. The presence of complement was critical for local production of cytokines (IFN-gamma and IL-10), chemokines (IP-10), and adhesion molecules (ICAM-1 and LECAM-1) during EAAU. Furthermore, intraocular complement activation, specifically iC3b production and engagement of complement receptor 3 (CR3), had a significant impact on disease activity in EAAU.
Conclusions: The study provided the novel finding that complement activation plays a central role in the pathogenesis of ocular autoimmunity and may serve as a potential target for therapeutic intervention.
Figures
Similar articles
-
Suppression of complement activation by recombinant Crry inhibits experimental autoimmune anterior uveitis (EAAU).Mol Immunol. 2010 Nov-Dec;48(1-3):231-9. doi: 10.1016/j.molimm.2010.08.006. Epub 2010 Sep 16. Mol Immunol. 2010. PMID: 20843553 Free PMC article.
-
Inhibition of complement alternative pathway suppresses experimental autoimmune anterior uveitis by modulating T cell responses.J Biol Chem. 2011 Mar 11;286(10):8472-8480. doi: 10.1074/jbc.M110.197616. Epub 2011 Jan 7. J Biol Chem. 2011. PMID: 21216963 Free PMC article.
-
Kinetics of cytokine production in experimental autoimmune anterior uveitis (EAAU).Curr Eye Res. 1998 Oct;17(10):955-61. doi: 10.1076/ceyr.17.10.955.5246. Curr Eye Res. 1998. PMID: 9788297
-
Intraocular diseases - anterior uveitis.Chem Immunol Allergy. 2007;92:213-220. doi: 10.1159/000099272. Chem Immunol Allergy. 2007. PMID: 17264497 Review.
-
What causes relapses of autoimmune diseases? The etiological role of autoreactive T cells.Autoimmun Rev. 2013 Sep;12(11):1070-5. doi: 10.1016/j.autrev.2013.04.001. Epub 2013 May 16. Autoimmun Rev. 2013. PMID: 23685277 Review.
Cited by
-
Tolerance to melanin-associated antigen in autoimmune uveitis is mediated by CD4+CD25+ T-regulatory cells.Am J Pathol. 2008 Nov;173(5):1440-54. doi: 10.2353/ajpath.2008.080150. Epub 2008 Oct 2. Am J Pathol. 2008. PMID: 18832572 Free PMC article.
-
Candidate Tear-Based Uveitis Biomarkers in Children with JIA Based on Arthritis Activity and Topical Corticosteroid Use.Ocul Immunol Inflamm. 2025 May;33(4):603-612. doi: 10.1080/09273948.2024.2428846. Epub 2024 Nov 25. Ocul Immunol Inflamm. 2025. PMID: 39586039
-
Copy number variations and gene polymorphisms of complement components in ocular Behcet's disease and Vogt-Koyanagi-Harada syndrome.Sci Rep. 2015 Aug 13;5:12989. doi: 10.1038/srep12989. Sci Rep. 2015. PMID: 26269006 Free PMC article.
-
Suppression of complement activation by recombinant Crry inhibits experimental autoimmune anterior uveitis (EAAU).Mol Immunol. 2010 Nov-Dec;48(1-3):231-9. doi: 10.1016/j.molimm.2010.08.006. Epub 2010 Sep 16. Mol Immunol. 2010. PMID: 20843553 Free PMC article.
-
As in Real Estate, Location Matters: Cellular Expression of Complement Varies Between Macular and Peripheral Regions of the Retina and Supporting Tissues.Front Immunol. 2022 Jun 15;13:895519. doi: 10.3389/fimmu.2022.895519. eCollection 2022. Front Immunol. 2022. PMID: 35784369 Free PMC article.
References
-
- Thomlinson S. Complement defense mechanisms. Curr Opin Immunol. 1993;5:83–89. - PubMed
-
- Dempsey PW, Allison MED, Akkaraju S, Goodnow CC, Fearon DT. C3d of complement as a molecular adjuvant: bridging innate and acquired immunity. Science. 1996;271:348–350. - PubMed
-
- Carroll MC. The complement system in regulation of adaptive immunity. Nat Immunol. 2004;5:981–986. - PubMed
-
- Pasinetti GM. Inflammatory mechanisms in neurodegeneration and Alzheimer’s disease: the role of the complement system. Neurobiol Aging. 1996;17:707–716. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
