

Natural killer cell behavior in lymph nodes revealed by static and real-time imaging
- PMID: 16505138
- PMCID: PMC2118232
- DOI: 10.1084/jem.20051474
Natural killer cell behavior in lymph nodes revealed by static and real-time imaging
Abstract
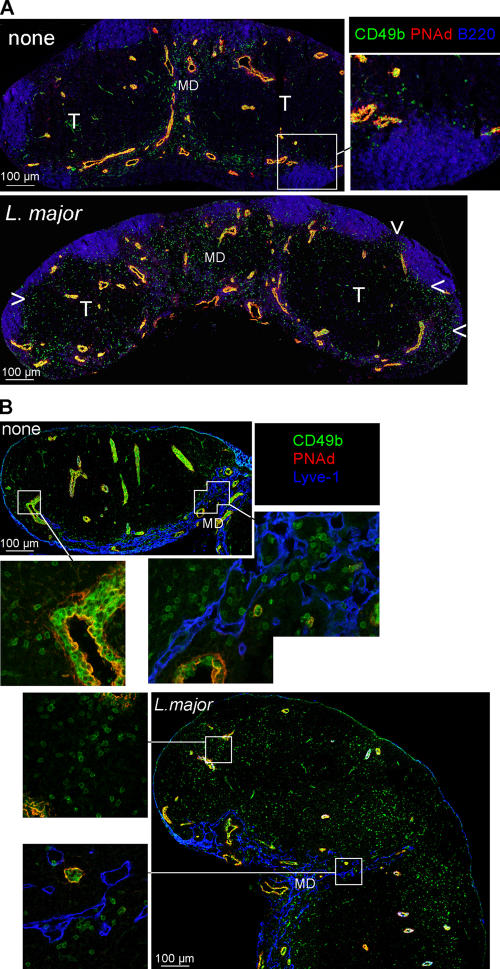
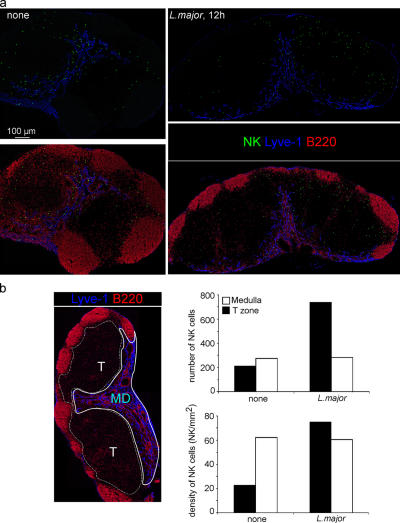
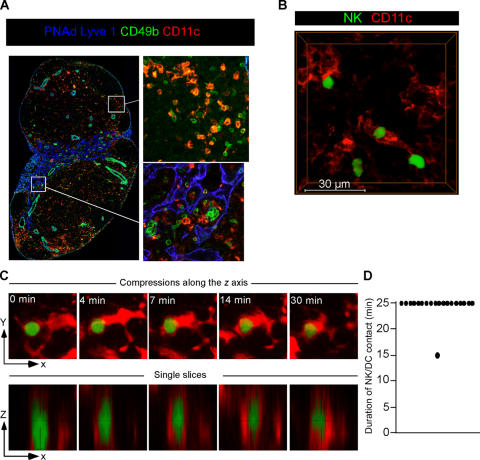
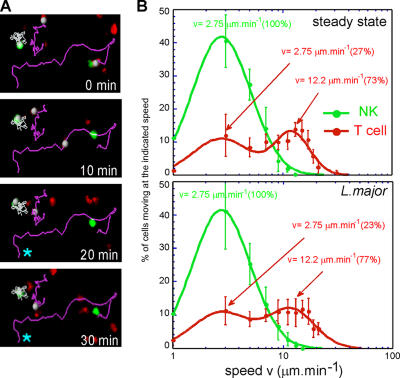
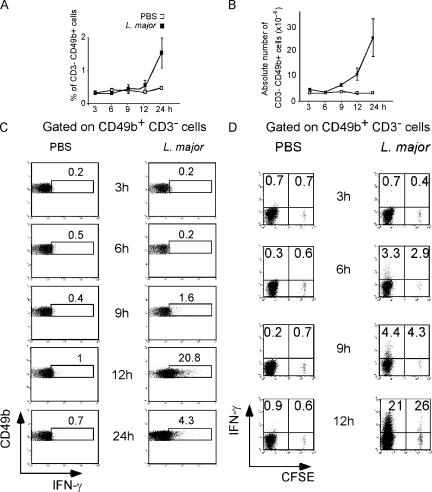
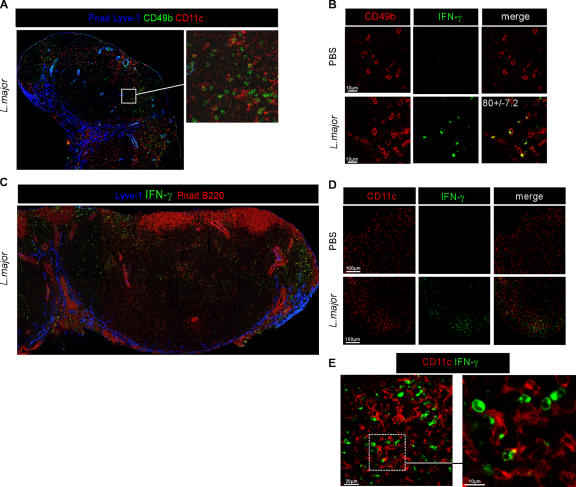
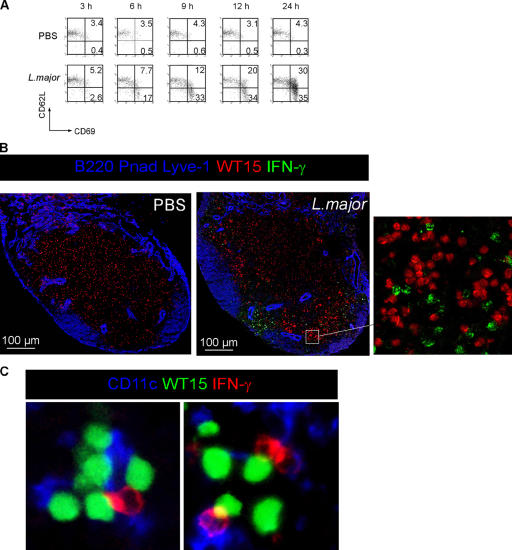
Natural killer (NK) cells promote dendritic cell (DC) maturation and influence T cell differentiation in vitro. To better understand the nature of the putative interactions among these cells in vivo during the early phases of an adaptive immune response, we have used immunohistochemical analysis and dynamic intravital imaging to study NK cell localization and behavior in lymph nodes (LNs) in the steady state and shortly after infection with Leishmania major. In the LNs of naive mice, NK cells reside in the medulla and the paracortex, where they closely associate with DCs. In contrast to T cells, intravital microscopy revealed that NK cells in the superficial regions of LNs were slowly motile and maintained their interactions with DCs over extended times in the presence or absence of immune-activating signals. L. major induced NK cells to secrete interferon-gamma and to be recruited to the paracortex, where concomitant CD4 T cell activation occurred. Therefore, NK cells form a reactive but low mobile network in a strategic area of the LN where they can receive inflammatory signals, interact with DCs, and regulate colocalized T cell responses.
Figures
References
-
- French, A.R., and W.M. Yokoyama. 2003. Natural killer cells and viral infections. Curr. Opin. Immunol. 15:45–51. - PubMed
-
- Yokoyama, W.M., S. Kim, and A.R. French. 2004. The dynamic life of natural killer cells. Annu. Rev. Immunol. 22:405–429. - PubMed
-
- Fernandez, N.C., A. Lozier, C. Flament, P. Ricciardi-Castagnoli, D. Bellet, M. Suter, M. Perricaudet, T. Tursz, E. Maraskovsky, and L. Zitvogel. 1999. Dendritic cells directly trigger NK cell functions: cross-talk relevant in innate anti-tumor immune responses in vivo. Nat. Med. 5:405–411. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
