

Access of extracellular cations to their binding sites in Na,K-ATPase: role of the second extracellular loop of the alpha subunit
- PMID: 16505152
- PMCID: PMC2151497
- DOI: 10.1085/jgp.200509418
Access of extracellular cations to their binding sites in Na,K-ATPase: role of the second extracellular loop of the alpha subunit
Abstract
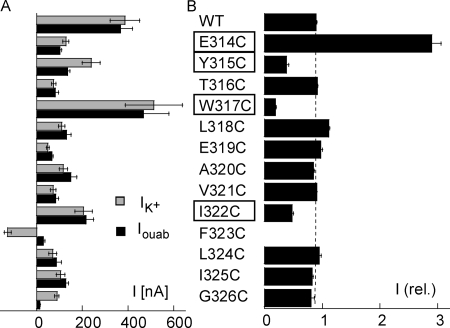
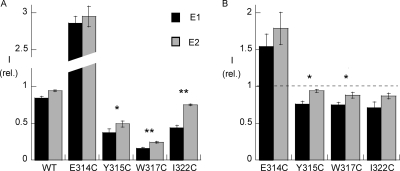
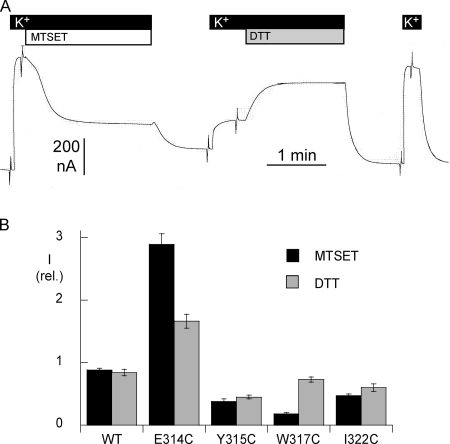
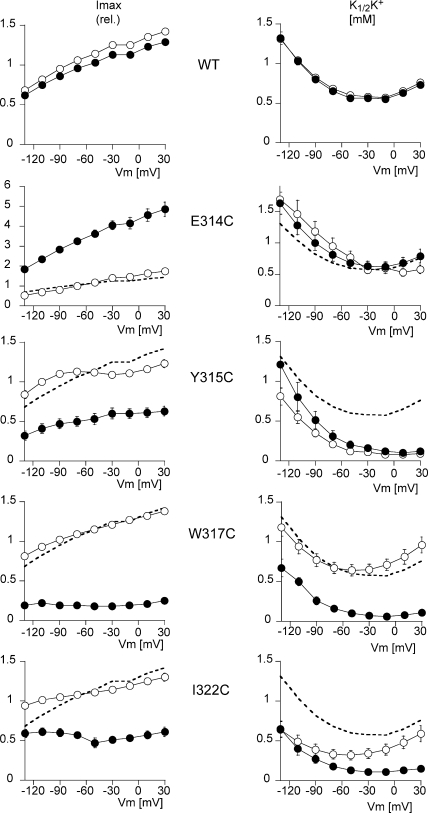
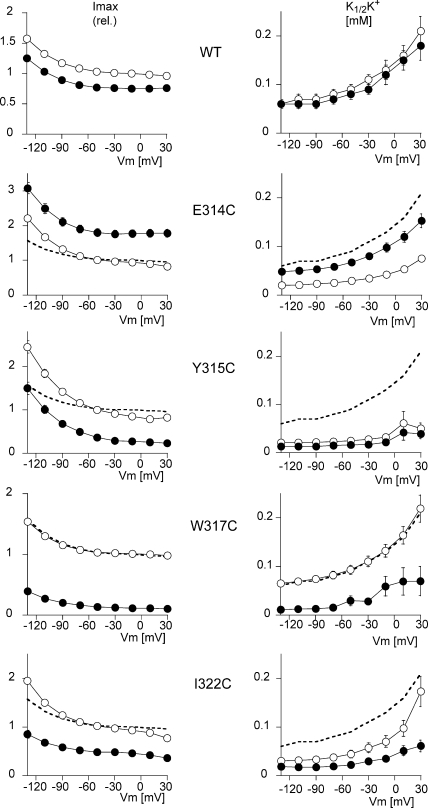
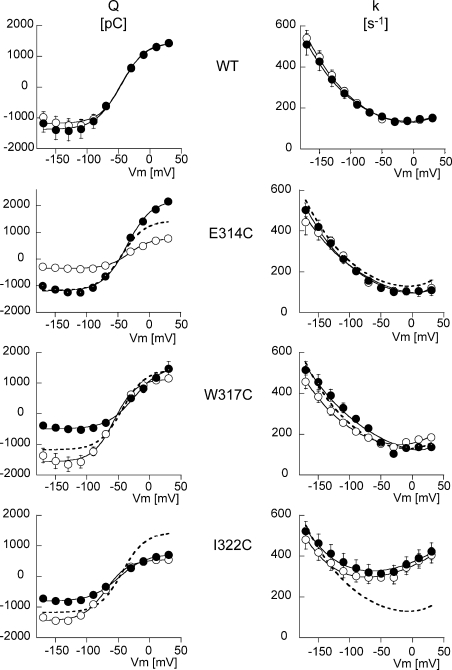
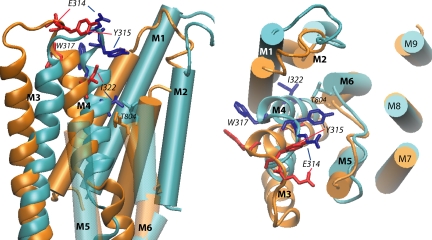
Na,K-ATPase, the main active transport system for monovalent cations in animal cells, is responsible for maintaining Na(+) and K(+) gradients across the plasma membrane. During its transport cycle it binds three cytoplasmic Na(+) ions and releases them on the extracellular side of the membrane, and then binds two extracellular K(+) ions and releases them into the cytoplasm. The fourth, fifth, and sixth transmembrane helices of the alpha subunit of Na,K-ATPase are known to be involved in Na(+) and K(+) binding sites, but the gating mechanisms that control the access of these ions to their binding sites are not yet fully understood. We have focused on the second extracellular loop linking transmembrane segments 3 and 4 and attempted to determine its role in gating. We replaced 13 residues of this loop in the rat alpha1 subunit, from E314 to G326, by cysteine, and then studied the function of these mutants using electrophysiological techniques. We analyzed the results using a structural model obtained by homology with SERCA, and ab initio calculations for the second extracellular loop. Four mutants were markedly modified by the sulfhydryl reagent MTSET, and we investigated them in detail. The substituted cysteines were more readily accessible to MTSET in the E1 conformation for the Y315C, W317C, and I322C mutants. Mutations or derivatization of the substituted cysteines in the second extracellular loop resulted in major increases in the apparent affinity for extracellular K(+), and this was associated with a reduction in the maximum activity. The changes produced by the E314C mutation were reversed by MTSET treatment. In the W317C and I322C mutants, MTSET also induced a moderate shift of the E1/E2 equilibrium towards the E1(Na) conformation under Na/Na exchange conditions. These findings indicate that the second extracellular loop must be functionally linked to the gating mechanism that controls the access of K(+) to its binding site.
Figures
References
-
- Apell, H.-J., and S.J.D. Karlish. 2001. Functional properties of Na,K-ATPase, and their structural implications, as detected with biophysical techniques. J. Membr. Biol. 180:1–9. - PubMed
-
- Artigas, P., N. Reyes, and D.C. Gadsby. 2005. Na,K-ATPase ion translocation pathway. J. Gen. Physiol. 126:4A (Abstr.).
-
- Brooks, B.R., R.E. Bruccoleri, B.D. Olafson, D.J. States, S. Swaminathan, and M. Karplus. 1983. Charmm-a program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 4:187–217.
-
- Canessa, C.M., J.-D. Horisberger, and B.C. Rossier. 1993. Mutation of a tyrosine in the H3-H4 ectodomain of Na,K-ATPase confers ouabain resistance. J. Biol. Chem. 268:17722–17726. - PubMed
