

Endocytic function of von Hippel-Lindau tumor suppressor protein regulates surface localization of fibroblast growth factor receptor 1 and cell motility
- PMID: 16505488
- PMCID: PMC2367321
- DOI: 10.1074/jbc.M511621200
Endocytic function of von Hippel-Lindau tumor suppressor protein regulates surface localization of fibroblast growth factor receptor 1 and cell motility
Abstract
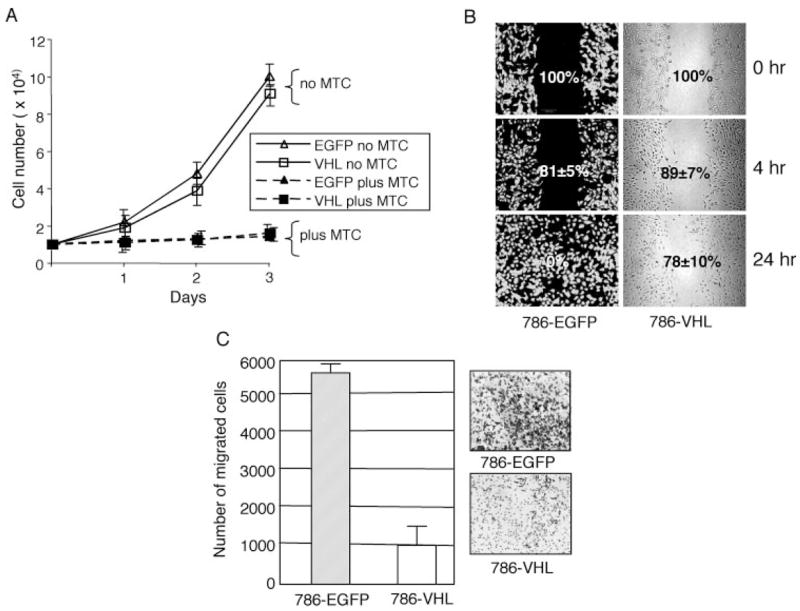
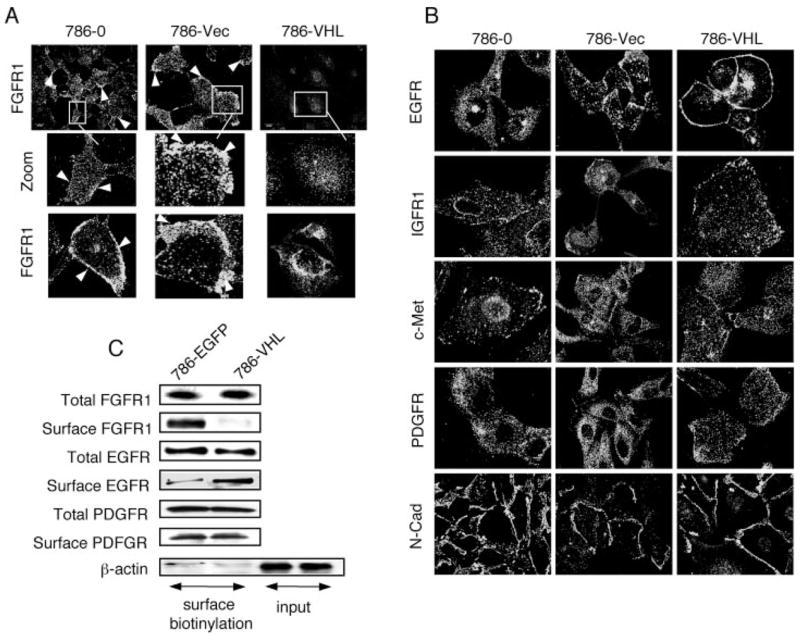
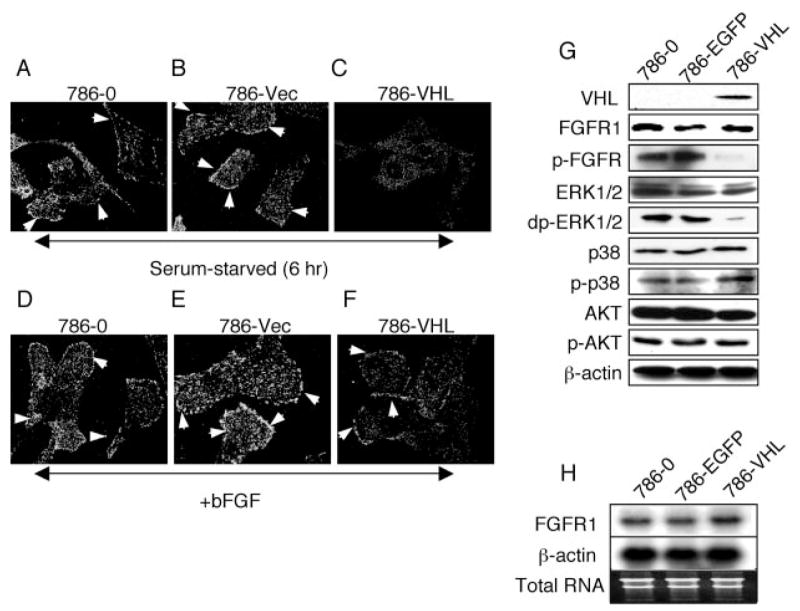
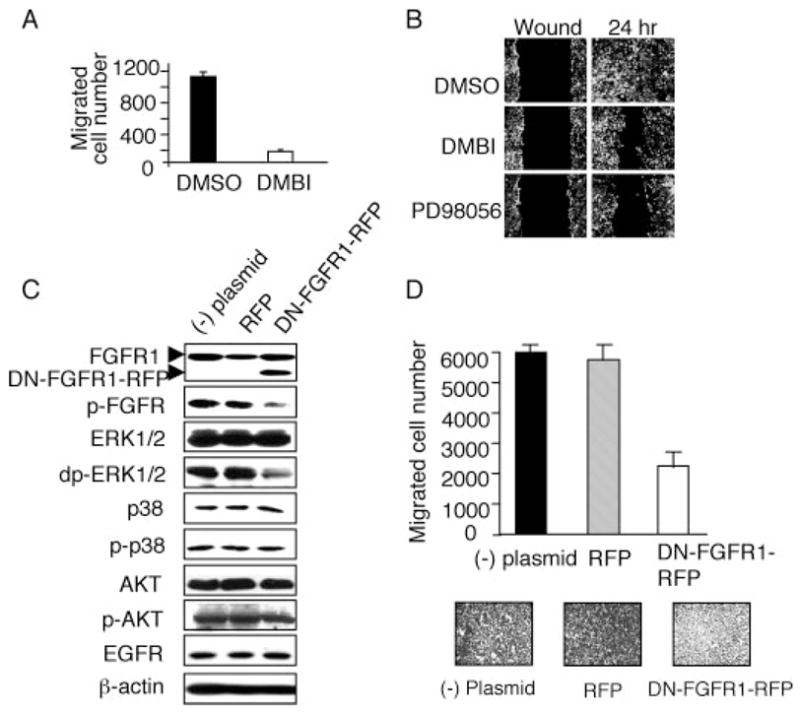
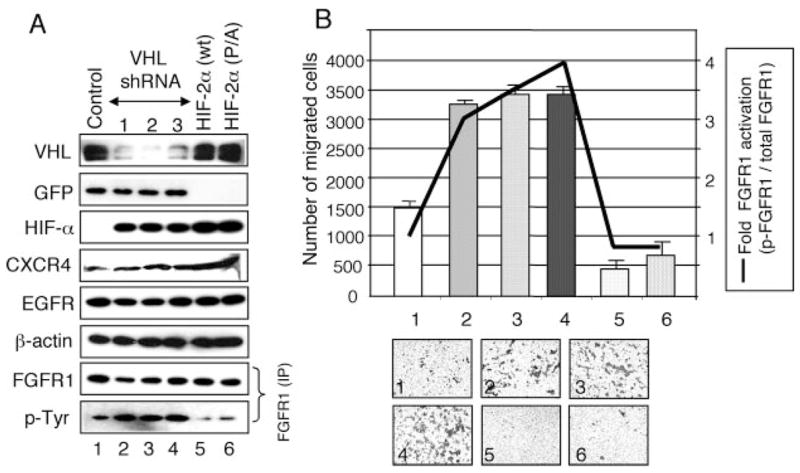
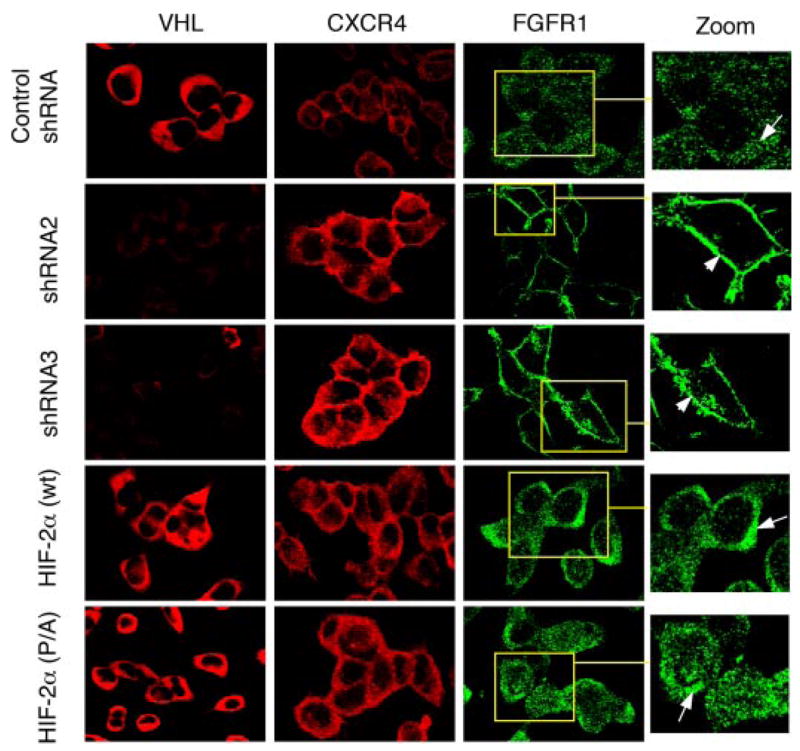
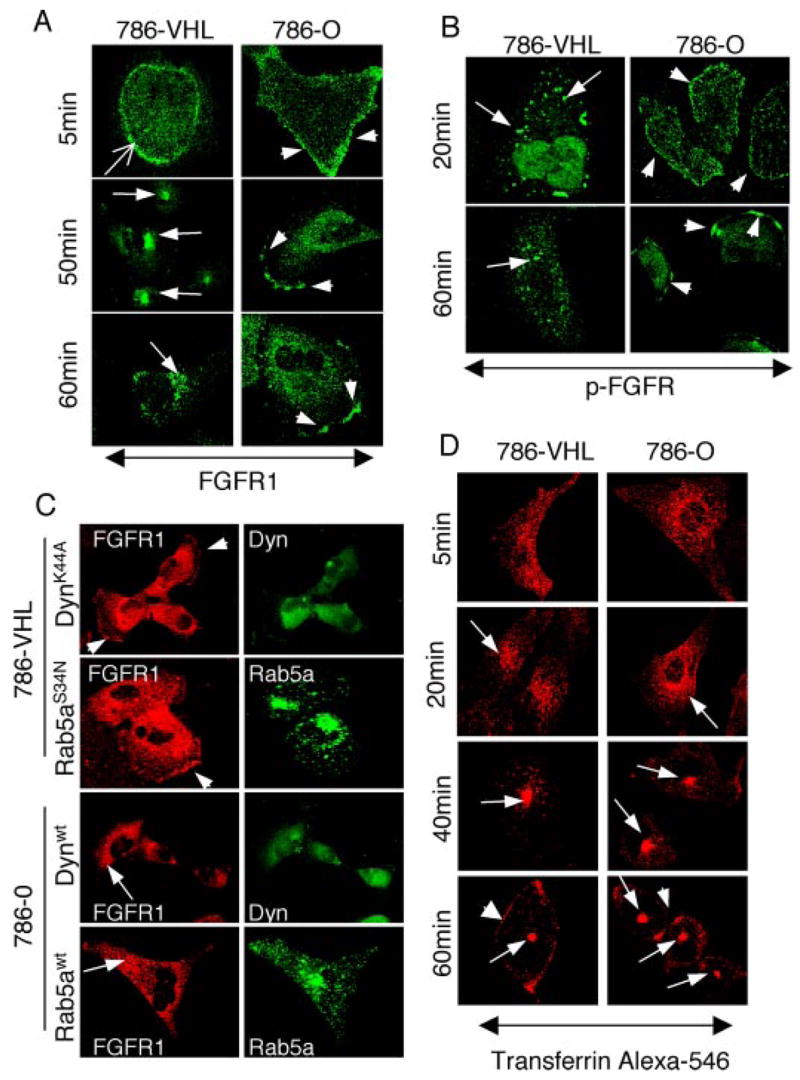
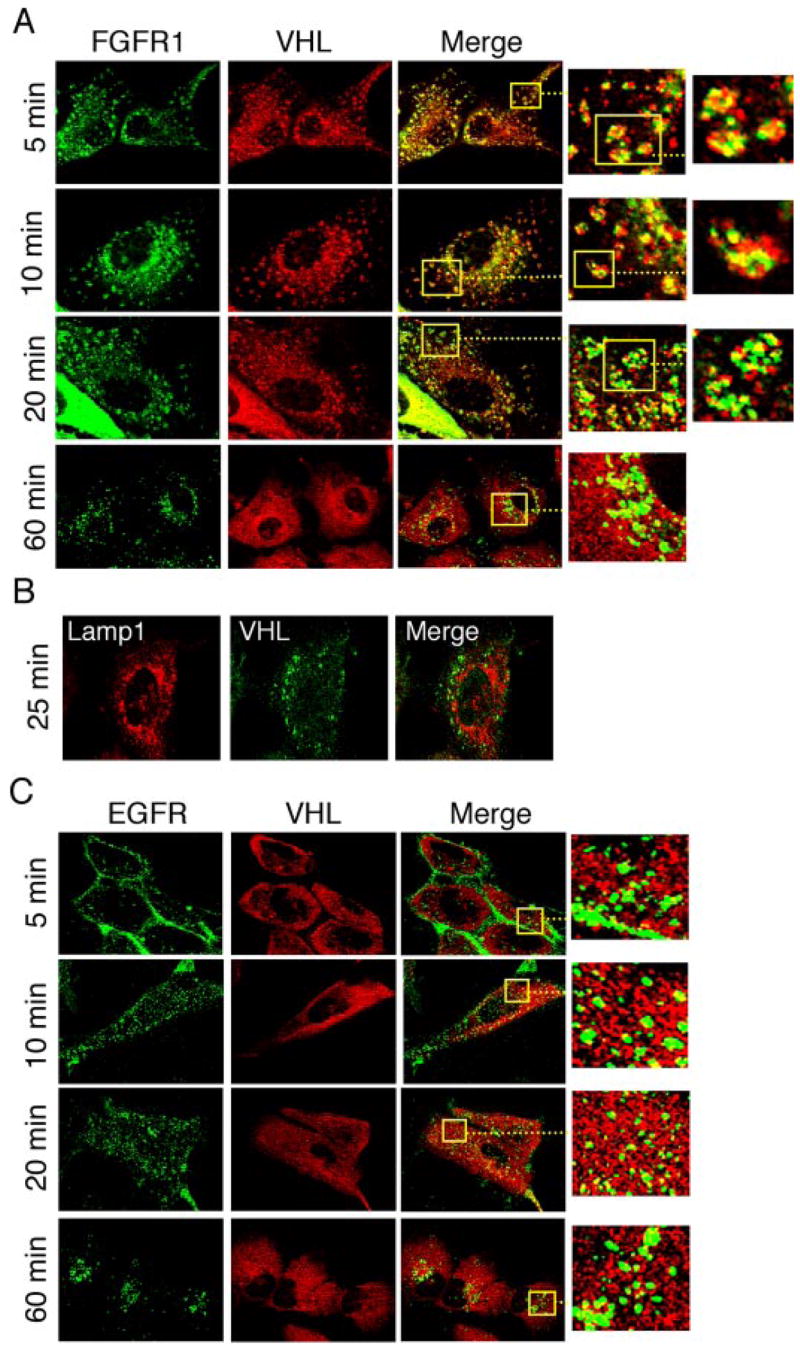
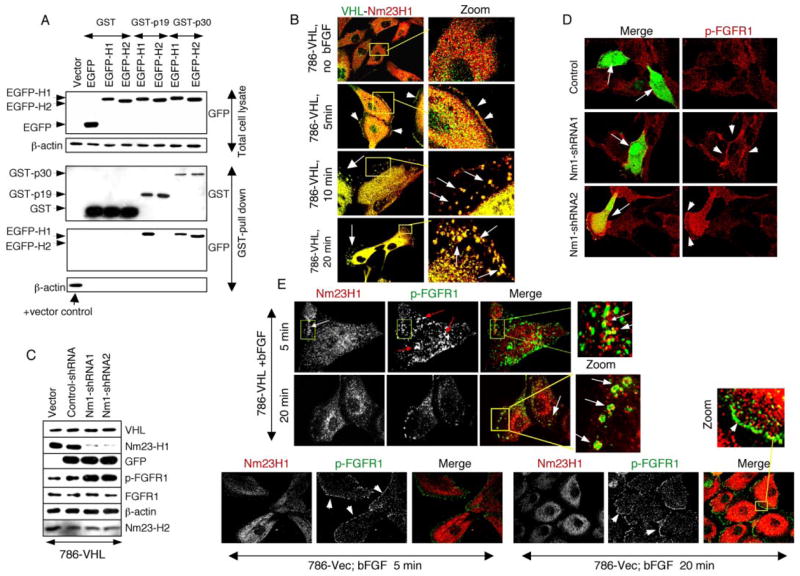
The tumor suppressor VHL (von Hippel-Lindau protein) serves as a negative regulator of hypoxia-inducible factor-alpha subunits. However, accumulated evidence indicates that VHL may play additional roles in other cellular functions. We report here a novel hypoxia-inducible factor-independent function of VHL in cell motility control via regulation of fibroblast growth factor receptor 1 (FGFR1) endocytosis. In VHL null tumor cells or VHL knock-down cells, FGFR1 internalization is defective, leading to surface accumulation and abnormal activation of FGFR1. The enhanced FGFR1 activity directly correlates with increased cell migration. VHL disease mutants, in two of the mutation hot spots favoring development of renal cell carcinoma, failed to rescue the above phenotype. Interestingly, surface accumulation of the chemotactic receptor appears to be selective in VHL mutant cells, since other surface proteins such as epidermal growth factor receptor, platelet-derived growth factor receptor, IGFR1, and c-Met are not affected. We demonstrate that 1) FGFR1 endocytosis is defective in the VHL mutant and is rescued by reexpression of wild-type VHL, 2) VHL is recruited to FGFR1-containing, but not EGFR-containing, endosomal vesicles, 3) VHL exhibits a functional relationship with Rab5a and dynamin 2 in FGFR1 internalization, and 4) the endocytic function of VHL is mediated through the metastasis suppressor Nm23, a protein known to regulate dynamin-dependent endocytosis.
Figures
Similar articles
-
Vascular endothelial growth factor-B and vascular endothelial growth factor-C expression in renal cell carcinomas: regulation by the von Hippel-Lindau gene and hypoxia.Cancer Res. 2001 Apr 1;61(7):3206-11. Cancer Res. 2001. PMID: 11306510
-
Endothelial function of von Hippel-Lindau tumor suppressor gene: control of fibroblast growth factor receptor signaling.Cancer Res. 2008 Jun 15;68(12):4649-57. doi: 10.1158/0008-5472.CAN-07-6003. Cancer Res. 2008. PMID: 18559510 Free PMC article.
-
Regulation of receptor for activated C kinase 1 protein by the von Hippel-Lindau tumor suppressor in IGF-I-induced renal carcinoma cell invasiveness.Oncogene. 2011 Feb 3;30(5):535-47. doi: 10.1038/onc.2010.427. Epub 2010 Sep 27. Oncogene. 2011. PMID: 20871634
-
[Tumor suppressor gene VHL, hypoxia inducible factor, and renal cell carcinoma].Zhonghua Bing Li Xue Za Zhi. 2006 Sep;35(9):562-4. Zhonghua Bing Li Xue Za Zhi. 2006. PMID: 17134554 Review. Chinese. No abstract available.
-
The von Hippel-Lindau tumor suppressor protein and clear cell renal carcinoma.Clin Cancer Res. 2007 Jan 15;13(2 Pt 2):680s-684s. doi: 10.1158/1078-0432.CCR-06-1865. Clin Cancer Res. 2007. PMID: 17255293 Review.
Cited by
-
Nm23-H1/NDP kinase folding intermediates and cancer: a hypothesis.J Bioenerg Biomembr. 2006 Aug;38(3-4):265-8. doi: 10.1007/s10863-006-9042-1. Epub 2006 Sep 1. J Bioenerg Biomembr. 2006. PMID: 16944300
-
Von Hippel-Lindau syndrome: molecular mechanisms of the disease.Clin Transl Oncol. 2010 Mar;12(3):160-5. doi: 10.1007/s12094-010-0485-9. Clin Transl Oncol. 2010. PMID: 20231120 Review.
-
Candidate tumor suppressor and pVHL partner Jade-1 binds and inhibits AKT in renal cell carcinoma.Cancer Res. 2013 Sep 1;73(17):5371-80. doi: 10.1158/0008-5472.CAN-12-4707. Epub 2013 Jul 1. Cancer Res. 2013. PMID: 23824745 Free PMC article.
-
Controlled Plasma Membrane Delivery of FGFR1 and Modulation of Signaling by a Novel Regulated Anterograde RTK Transport Pathway.Cancers (Basel). 2023 Dec 14;15(24):5837. doi: 10.3390/cancers15245837. Cancers (Basel). 2023. PMID: 38136383 Free PMC article.
-
The Role of VHL in the Development of von Hippel-Lindau Disease and Erythrocytosis.Genes (Basel). 2022 Feb 17;13(2):362. doi: 10.3390/genes13020362. Genes (Basel). 2022. PMID: 35205407 Free PMC article. Review.
References
-
- Lonser RR, Glenn GM, Walther M, Chew EY, Libutti SK, Linehan WM, Oldfield EH. Lancet. 2003;361:2059–2067. - PubMed
-
- Kim WY, Kaelin WG. J Clin Oncol. 2004;22:4991–5004. - PubMed
-
- Latif F, Tory K, Gnarra J, Yao M, Duh FM, Orcutt ML, Stackhouse T, Kuzmin I, Modi W, Geil L. Science. 1993;260:1317–1320. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
