

c-Myb is an essential downstream target for homeobox-mediated transformation of hematopoietic cells
- PMID: 16507773
- PMCID: PMC1895838
- DOI: 10.1182/blood-2005-12-5014
c-Myb is an essential downstream target for homeobox-mediated transformation of hematopoietic cells
Abstract
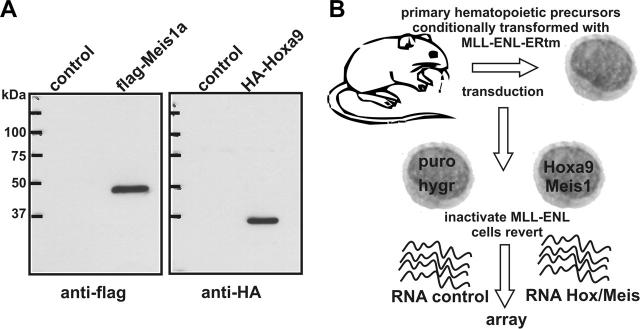
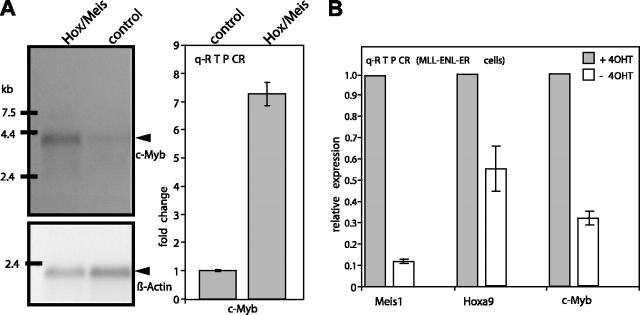
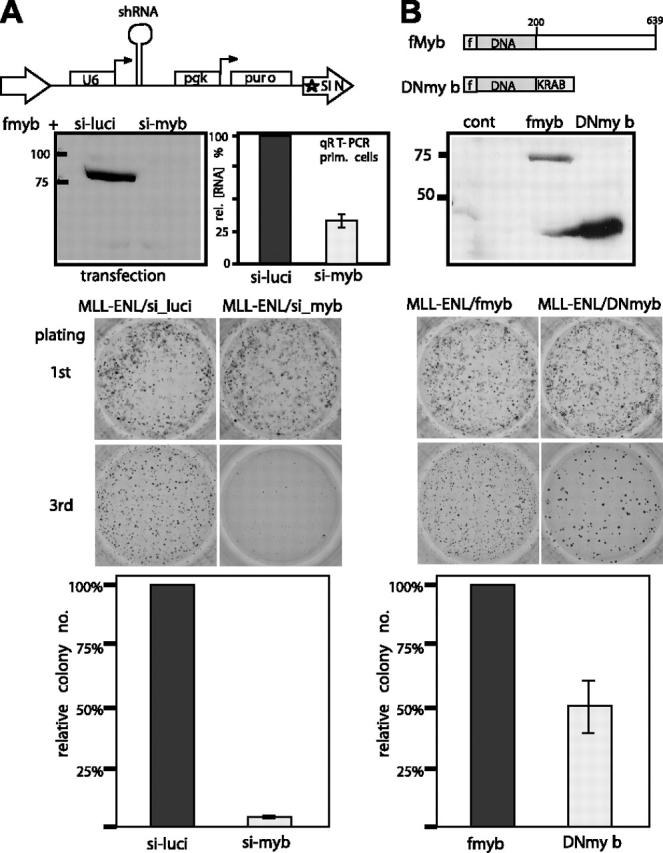
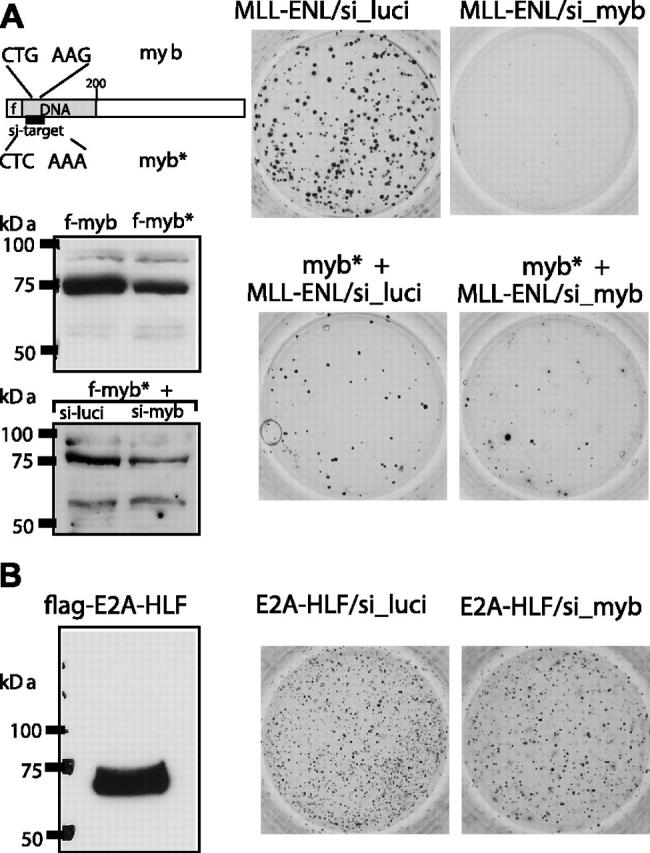
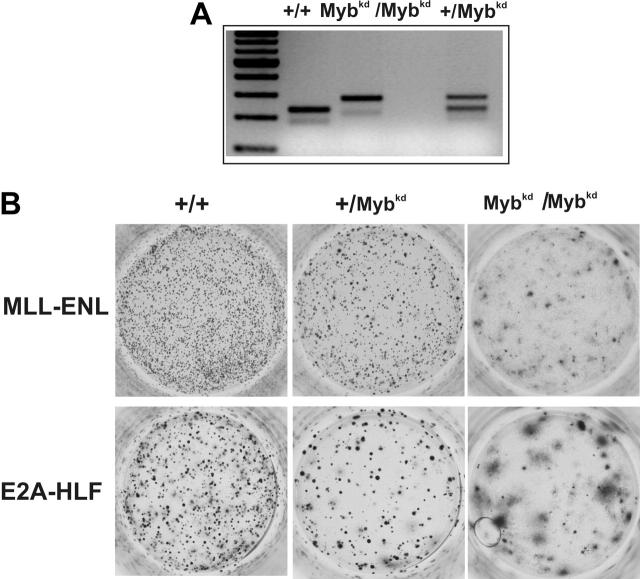
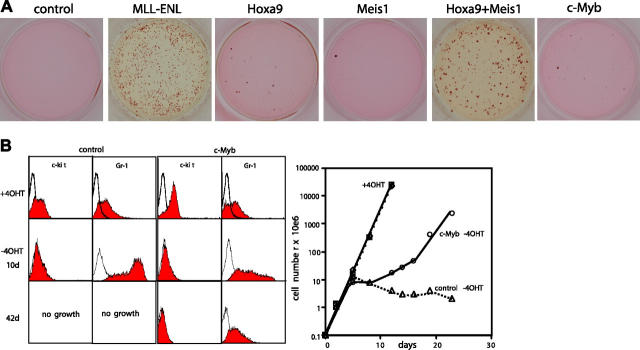
Abdominal-type HoxA genes in combination with Meis1 are well-documented on-cogenes in various leukemias but it is unclear how they exert their transforming function. Here we used a system of conditional transformation by an inducible mixed lineage leukemia-eleven-nineteen leukemia (MLL-ENL) oncoprotein to overexpress Hoxa9 and Meis1 in primary hematopoietic cells. Arrays identified c-Myb and a c-Myb target (Gstm1) among the genes with the strongest response to Hoxa9/Meis1. c-Myb overexpression was verified by Northern blot and quantitative reverse transcription-polymerase chain reaction (RT-PCR). Also MLL-ENL activated c-Myb through up-regulation of Hoxa9 and Meis1. Consequently, short-term suppression of c-Myb by small inhibitory RNA (siRNA) efficiently inhibited transformation by MLL-ENL but did not impair transformation by transcription factor E2A-hepatic leukemia factor (E2A-HLF). The anti c-Myb siRNA effect was abrogated by coexpression of a c-Myb derivative with a mutated siRNA target site. The introduction of a dominant-negative c-Myb mutant had a similar but weaker effect on MLL-ENL-mediated transformation. Hematopoietic precursors from mice homozygous for a hypo-morphic c-Myb allele were more severely affected and could be transformed neither by MLL-ENL nor by E2A-HLF. Ectopic expression of c-Myb induced a differentiation block but c-Myb alone was not transforming in a replating assay similar to Hoxa9/Meis1. These results suggest that c-Myb is essential but not sufficient for Hoxa9/Meis1 mediated transformation.
Figures
References
-
- Abramovich C, Humphries RK. Hox regulation of normal and leukemic hematopoietic stem cells. Curr Opin Hematol. 2005;12: 210-216. - PubMed
-
- Abramovich C, Pineault N, Ohta H, Humphries RK. Hox genes: from leukemia to hematopoietic stem cell expansion. Ann N Y Acad Sci. 2005;1044: 109-116. - PubMed
-
- Pineault N, Helgason CD, Lawrence HJ, Humphries RK. Differential expression of Hox, Meis1, and Pbx1 genes in primitive cells throughout murine hematopoietic ontogeny. Exp Hematol. 2002;30: 49-57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
