

Regulation and role of Raf-1/B-Raf heterodimerization
- PMID: 16508002
- PMCID: PMC1430271
- DOI: 10.1128/MCB.26.6.2262-2272.2006
Regulation and role of Raf-1/B-Raf heterodimerization
Abstract
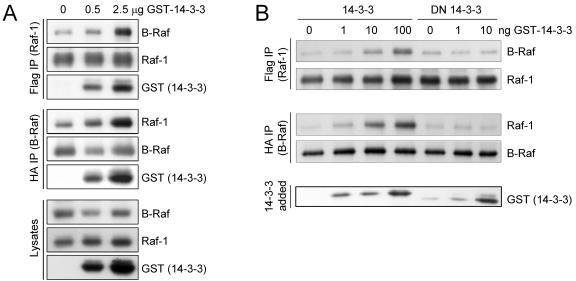
The Ras-Raf-MEK-extracellular signal-regulated kinase (ERK) pathway participates in the control of many fundamental cellular processes including proliferation, survival, and differentiation. The pathway is deregulated in up to 30% of human cancers, often due to mutations in Ras and the B-Raf isoform. Raf-1 and B-Raf can form heterodimers, and this may be important for cellular transformation. Here, we have analyzed the biochemical and biological properties of Raf-1/B-Raf heterodimers. Isolated Raf-1/B-Raf heterodimers possessed a highly increased kinase activity compared to the respective homodimers or monomers. Heterodimers between wild-type Raf-1 and B-Raf mutants with low or no kinase activity still displayed elevated kinase activity, as did heterodimers between wild-type B-Raf and kinase-negative Raf-1. In contrast, heterodimers containing both kinase-negative Raf-1 and kinase-negative B-Raf were completely inactive, suggesting that the kinase activity of the heterodimer specifically originates from Raf and that either kinase-competent Raf isoform is sufficient to confer high catalytic activity to the heterodimer. In cell lines, Raf-1/B-Raf heterodimers were found at low levels. Heterodimerization was enhanced by 14-3-3 proteins and by mitogens independently of ERK. However, ERK-induced phosphorylation of B-Raf on T753 promoted the disassembly of Raf heterodimers, and the mutation of T753 prolonged growth factor-induced heterodimerization. The B-Raf T753A mutant enhanced differentiation of PC12 cells, which was previously shown to be dependent on sustained ERK signaling. Fine mapping of the interaction sites by peptide arrays suggested a complex mode of interaction involving multiple contact sites with a main Raf-1 binding site in B-Raf encompassing T753. In summary, our data suggest that Raf-1/B-Raf heterodimerization occurs as part of the physiological activation process and that the heterodimer has distinct biochemical properties that may be important for the regulation of some biological processes.
Figures





References
-
- Abraham, D., K. Podar, M. Pacher, M. Kubicek, N. Welzel, B. A. Hemmings, S. M. Dilworth, H. Mischak, W. Kolch, and M. Baccarini. 2000. Raf-1-associated protein phosphatase 2A as a positive regulator of kinase activation. J. Biol. Chem. 275:22300-22304. - PubMed
-
- Alessi, D. R., P. Cohen, A. Ashworth, S. Cowley, S. J. Leevers, and C. J. Marshall. 1995. Assay and expression of mitogen-activated protein kinase, MAP kinase kinase, and Raf, p. 279-289. In W. E. Balch, C. J. Der, and A. Hall (ed.), Methods in enzymology. Academic Press, London, United Kingdom. - PubMed
-
- Brummer, T., H. Naegele, M. Reth, and Y. Misawa. 2003. Identification of novel ERK-mediated feedback phosphorylation sites at the C-terminus of B-Raf. Oncogene 22:8823-8834. - PubMed
-
- Chong, H., H. G. Vikis, and K. L. Guan. 2003. Mechanisms of regulating the Raf kinase family. Cell. Signal. 15:463-469. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous