

Characterization of different isoforms of the HIF prolyl hydroxylase PHD1 generated by alternative initiation
- PMID: 16509823
- PMCID: PMC1479752
- DOI: 10.1042/BJ20051996
Characterization of different isoforms of the HIF prolyl hydroxylase PHD1 generated by alternative initiation
Abstract
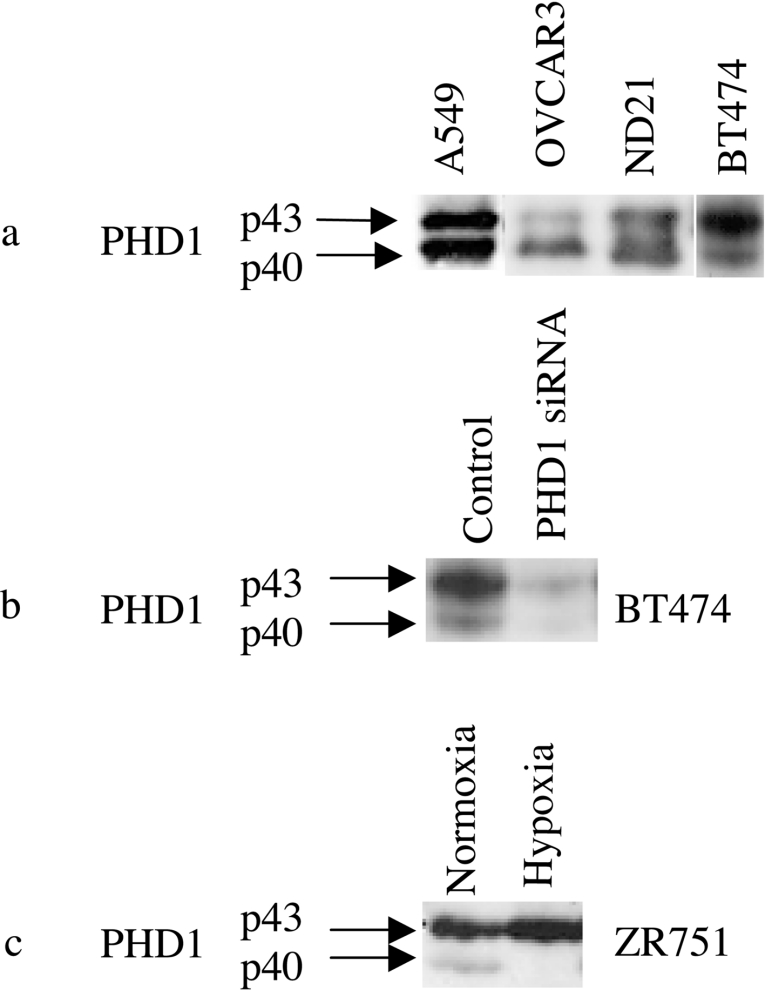
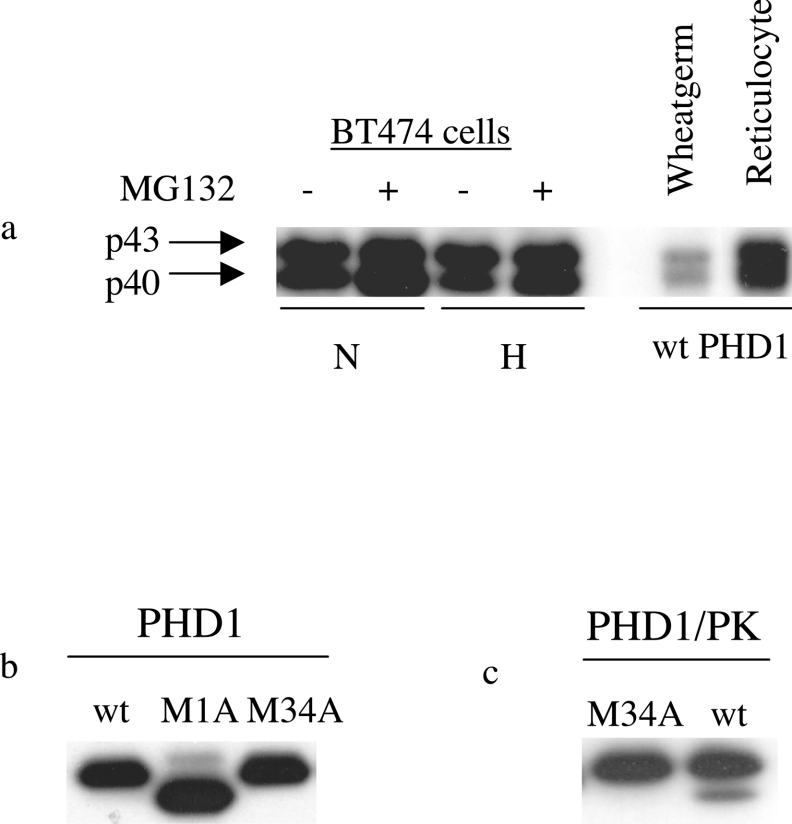
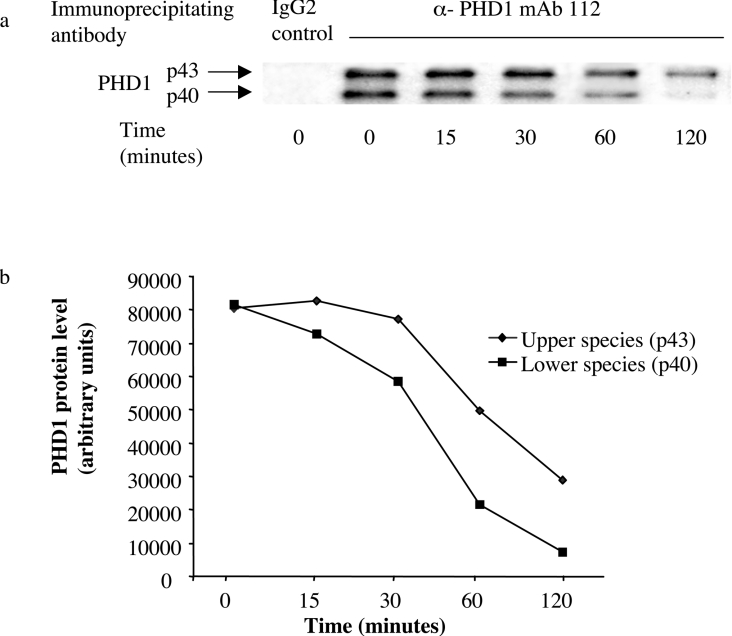
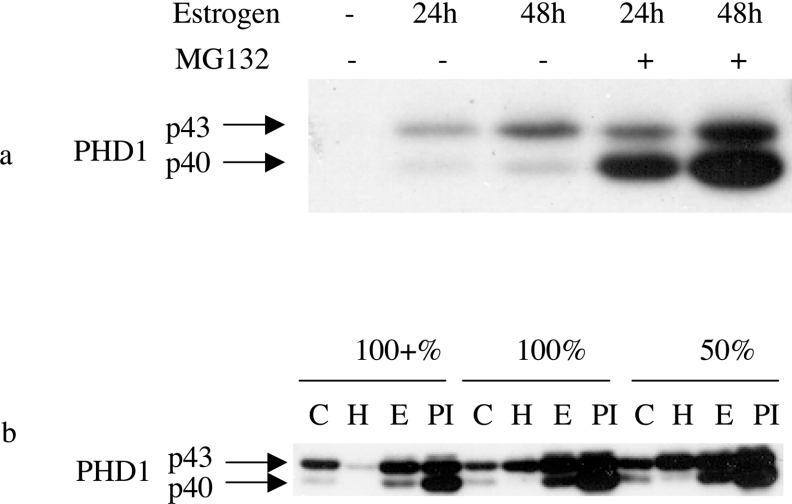
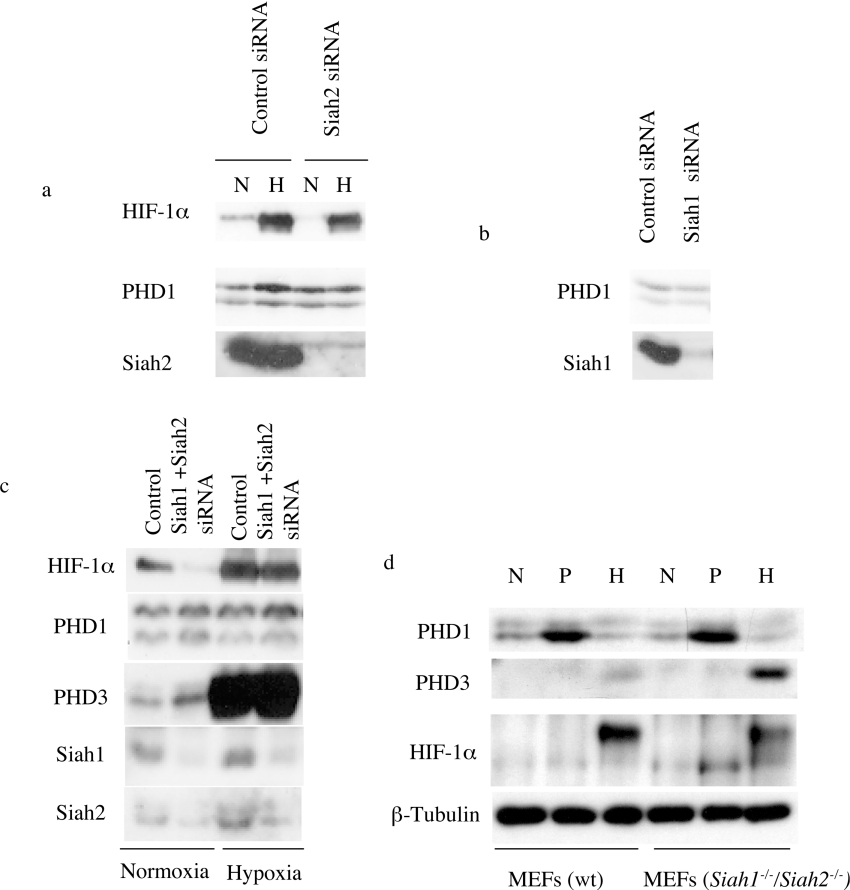
The heterodimeric transcription factor HIF (hypoxia-inducible factor) is central to the regulation of gene expression by oxygen. Three oxygen-dependent prolyl hydroxylase enzymes [PHD1 (prolyl hydroxylase domain 1), PHD2 and PHD3] control the abundance of HIF. In the presence of oxygen, they hydroxylate specific proline residues in HIF-alpha, allowing recognition by pVHL (von Hippel-Lindau protein) and subsequent ubiquitylation and proteasomal destruction. The precise roles and regulation of these enzymes are therefore of particular importance in understanding the physiological and pathological responses to hypoxia. In the present study, we define the existence of two species of PHD1 and provide evidence that they are generated by alternative translational initiation. We demonstrate that these alternative forms are both biologically active with similar HIF prolyl hydroxylase activity but that they differ in their responses to oestrogen, cell confluence and proteasomal inhibition. We show that the two PHD1 species are subject to proteolytic regulation but differ markedly in their protein stability. Though each isoform has the potential to interact with members of the Siah (seven in absentia homologue) ubiquitin ligase family, genetic studies indicated that other proteolytic mechanisms are responsible for control of stability under the conditions examined. The data define the existence of a further level of control in the pathway that regulates cellular responses to hypoxia.
Figures
References
-
- Kaelin W. G. J. How oxygen makes its presence felt. Genes Dev. 2002;16:1441–1445. - PubMed
-
- Wang G. L., Semenza G. L. Purification and characterisation of hypoxiainducible factor 1. J. Biol. Chem. 1995;270:1230–1237. - PubMed
-
- Maxwell P. H., Wiesener M. S., Chang G.-W., Clifford S. C., Vaux E. C., Cockman M. E., Wykoff C. C., Pugh C. W., Maher E. R., Ratcliffe P. J. The tumour suppressor protein VHL targets hypoxia-inducible factors for oxygen-dependent proteolysis. Nature (London) 1999;399:271–275. - PubMed
-
- Ivan M., Kondo K., Yang H., Kim W., Valiando J., Ohh M., Salic A., Asara J. M., Lane W. S., Kaelin W. G. J. HIFα targeted for VHL-mediated destruction by proline hydroxylation: implications for O2 sensing. Science. 2001;292:464–468. - PubMed
-
- Jaakkola P., Mole D. R., Tian Y.-M., Wilson M. I., Gielbert J., Gaskell S. J., Kriegsheim A. V., Hebestreit H. F., Mukherji M., Schofield C. J., et al. Targeting of HIF-α to the von Hippel-Lindau ubiquitylation complex by O2-regulated prolyl hydroxylation. Science. 2001;292:468–472. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases